







前言
休眠是由环境因素和内部分子机制通过复杂的相互作用进行严格调控的过程。了解休眠对于预测种群动态和探索不利生存条件下的适应机制至关重要。在不利的环境触发下,水母Aurelia coerulea 水螅态会形成休眠的关键标志——足囊,当恢复有利环境时,足囊发生外囊化,使水螅态发育并随后产生新的水母。A. coerulea的这种能力为研究休眠形成的进化机制提供了绝佳的机会。
2023年12月,中国科学院烟台海岸带研究所董志军团队在Molecular Ecology (IF:4.90)上发表题为“Environmental and molecular regulation of diapause formation in a scyphozoan jellyfish”的研究论文。该研究使用月水母 A. coerulea作为模式生物来研究温度和食物供应对休眠足囊形成的影响,并利用BD Rhapsody单细胞 RNA 测序技术探索了休眠形成的分子调控机制。
研究结果
温度升高和饥饿加速足囊形成
作者建立了8个具有两个环境因素 (温度、食物供应)的实验组来评估对足囊形成的影响。S 及 MS 分别代表饥饿和轻度饥饿处理,同时分别与4个温度处理(9 ℃、22 ℃、19 ℃和 4℃)组合,使用大小相似且情况良好的水螅态进行试验。经过 30 天的孵育期,NPP(非足囊形成性水螅态)和 PP(足囊形成性水螅态)之间观察到显着的形态差异。PP的触手收缩,花萼变细,茎伸长成透明的管状结构,每个管状结构中包裹着一个白色的、絮状的、球形的足囊。随后,PP 的茎收缩,水螅态重新定位,将球形足囊留在原来的附着位点。新形成的足囊形态总体呈球形,直径约为200 μm,外观柔软、透明、呈淡黄色。在不同的处理组中,观察到单个水螅态的出芽率和足囊形成率存在显著差异。MS19组的出芽率最高且足囊形成率最高。气温的升高和食物供应的减少与足囊形成率的增加呈正相关。值得注意的是,在4 ℃下,饥饿处理组和轻度饥饿处理组的水螅态均不形成足囊。
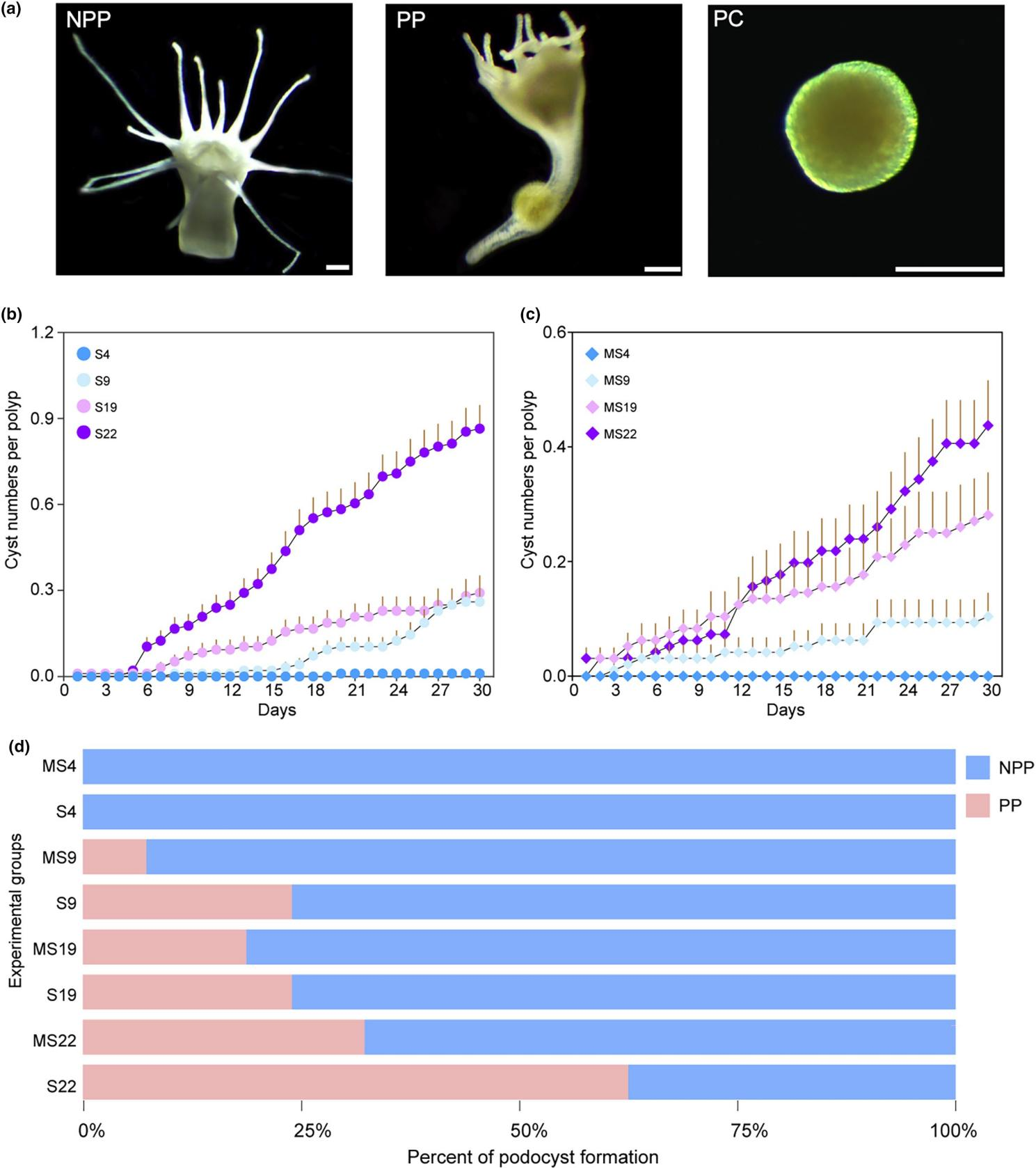
图 1 水螅态A. coerulea 在 30 天孵育期内不同处理条件下足囊的形成
NPP 与 PP 细胞图谱对比分析
为了系统地确定足囊形成过程中基因表达模式的动态变化,作者制备了NPP(非足囊形成性水螅态)和 PP(足囊形成性水螅态)的单细胞悬浮液。通过细胞群组中特定上调的基因特征结合已知Aurelia 标记基因或其他分类群中同源基因的数据,将细胞分为8个类型:表皮/肌肉细胞、胃真皮细胞、感觉毛细胞、刺细胞、免疫细胞、腺细胞、神经细胞、分化细胞。在对 NPP 和 PP的对比分析中,观察到细胞组成发生显著变化。表皮/肌肉细胞在 PP 中显著高于 NPP,但 PP 中的胃真皮细胞的比例显著低于 NPP,感觉毛细胞,刺细胞,免疫细胞,腺细胞,神经细胞及分化细胞的比例在两组间没有显著差异。
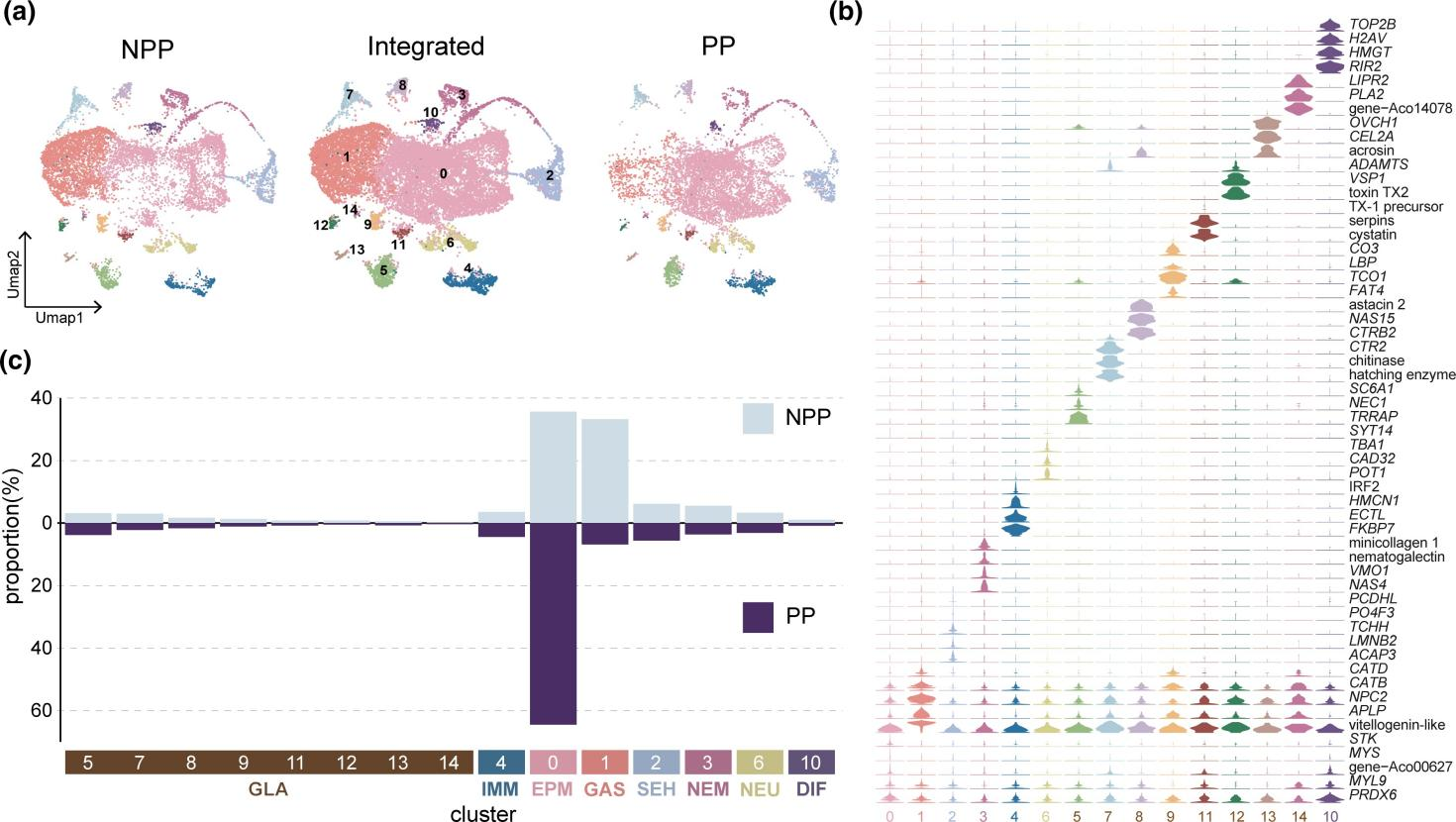
图 2 NPP 和 PP 细胞图谱比较
NPP 和 PP 之间基因表达模式的变化
在本研究中,作者发现在足囊的形成过程中,多种细胞类型的特定基因表达发生显著变化。NPP 和 PP 的刺细胞,免疫细胞,神经细胞和分化细胞的表达谱存在明显差异。相比之下,其他细胞类型之间基因表达的差异相对较小。DEG(差异表达基因)分析进一步表明所有细胞类型的基因表达存在不同程度的差异。刺细胞、腺细胞和免疫细胞在 NPP 和 PP 之间表现出较高数量的 DEG,计数分别为 38、33 和 21。在相同的细胞类型中,DEGs 在所有表达基因中所占的比例明显更高,刺细胞为 2.708%,腺细胞为 2.048%,免疫细胞为1.590%。在 PP 的大多数细胞类型中,HSC70、HSP90a、AMPO、JUN 和 CEBPB 等基因的表达显著上调,而 NPC2、VIT2 和 IF2B 的表达显著下调。在 PP 的刺细胞中,包括 NAS4、NAS15、PA23 和 MAC/穿孔素结构域在内的毒素相关基因显著下调。在PP的神经细胞中,信号转导相关基因的表达,如TBA1和AGTWamide,显著下调。
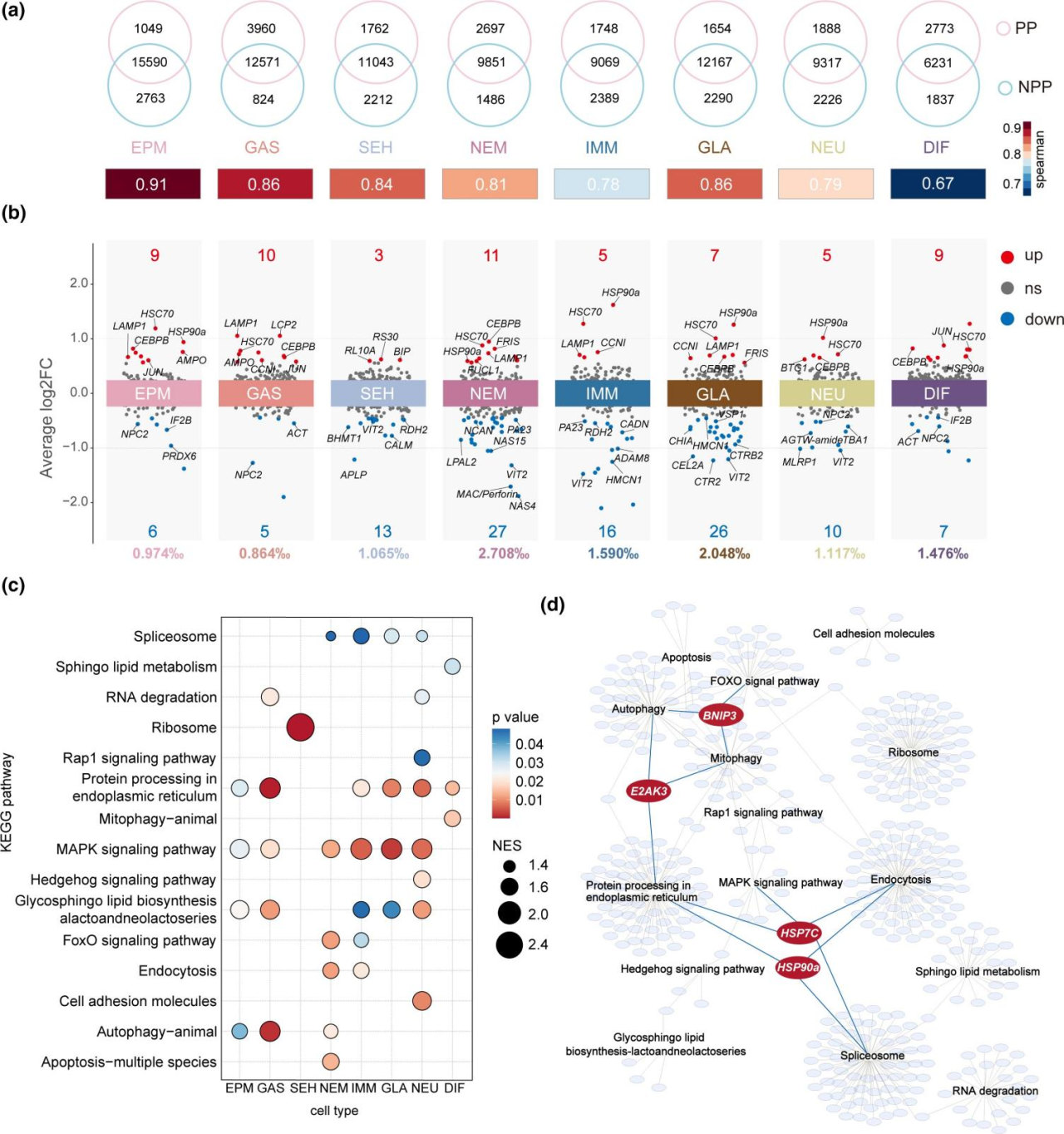
图 3 足囊形成性水螅态和非足囊形成性水螅态的基因表达谱比较
NPP 和 PP 之间转录因子的变化
为了探索转录因子TF 的表达与足囊形成之间的关系,作者对 NPP 和 PP 中不同细胞类型的 TF 动态进行了全面分析。作者鉴定了 212 个 TF 基因,它们在 NPP 和 PP 之间具有共同的 GO 注释。其中,14 个TF基因在 PP 中独特表达,而 3 个 TF 基因(HXB2、HM17和HME2B)是 NPP 特有的。例如,HOXD1 独特地表达于 PP 的表皮/肌肉细胞中,而 TEF3 仅在 PP 的刺细胞中表达。OPTIX 在 PP 的表皮/肌肉、胃真皮和感觉毛细胞中特异性表达。BARHL1 在 PP 的表皮/肌肉、感觉毛和腺细胞中表现出特异性。HOX1 和 RUNX1 在 PP 中分别为表皮/肌肉细胞和感觉毛细胞所独有。COE1 在 PP 的表皮/肌肉细胞和线母细胞中特异性表达。此外,包括 MYC、CEBPB、JUN、FOXO 和 ASCL4 在内的几个 TF 基因在 PP 中的表达显着高于 NPP。
休眠足囊的细胞图谱
为了阐明足囊的细胞组成和基因转录动态,作者从 MS22 组中分离出 20 个足囊并利用了基于微孔原理的BD Rhapsody单细胞多组学平台对单细胞进行分离捕获。根据不同的基因表达谱,作者将细胞分为六类。Cluster 0 主要表达核糖体基因,包括 RPL30、RPL35、RPL36、RPLP2 和铁蛋白。GO分析显示,Cluster 0富含与核糖体、翻译延伸和RNA结合相关的功能。这表明 Cluster 0 可能参与蛋白质合成。Cluster 1 表现出 JUN、HLF、ATF4 和 BTG2 的高表达。JUN 和 HLF 因其在细胞生长、分化和对外部刺激的反应中的作用而闻名,ATF4 与氨基酸代谢和细胞对氧化应激的反应有关,而 BTG2 与细胞周期调节和生长控制相关。GO分析表明, Cluster 1富含脂质转运、GTPase活性和ATPase活性,表明这些细胞可能参与细胞分化和代谢应激反应。使用标记 IRF1、ERG1 和 LBP 将 Cluster 2识别为免疫细胞。Cluster 3 特异性表达 POTEJ、HSC70 和 HSP90a。HSC70 和 HSP90a 对于细胞应激反应至关重要,有助于蛋白质折叠、稳定性和降解。这些基因在 Cluster 3 中的独特表达表明该Cluster可能在细胞应激反应中具有特殊的作用。Cluster 4 特异性表达 GAT1、CFA65 和 DNAAF5,并被鉴定为神经细胞。根据小胶原蛋白1、线半乳糖凝集素、NAS4 和 DKK3 的特异性表达,Cluster 5 被鉴定为刺细胞。
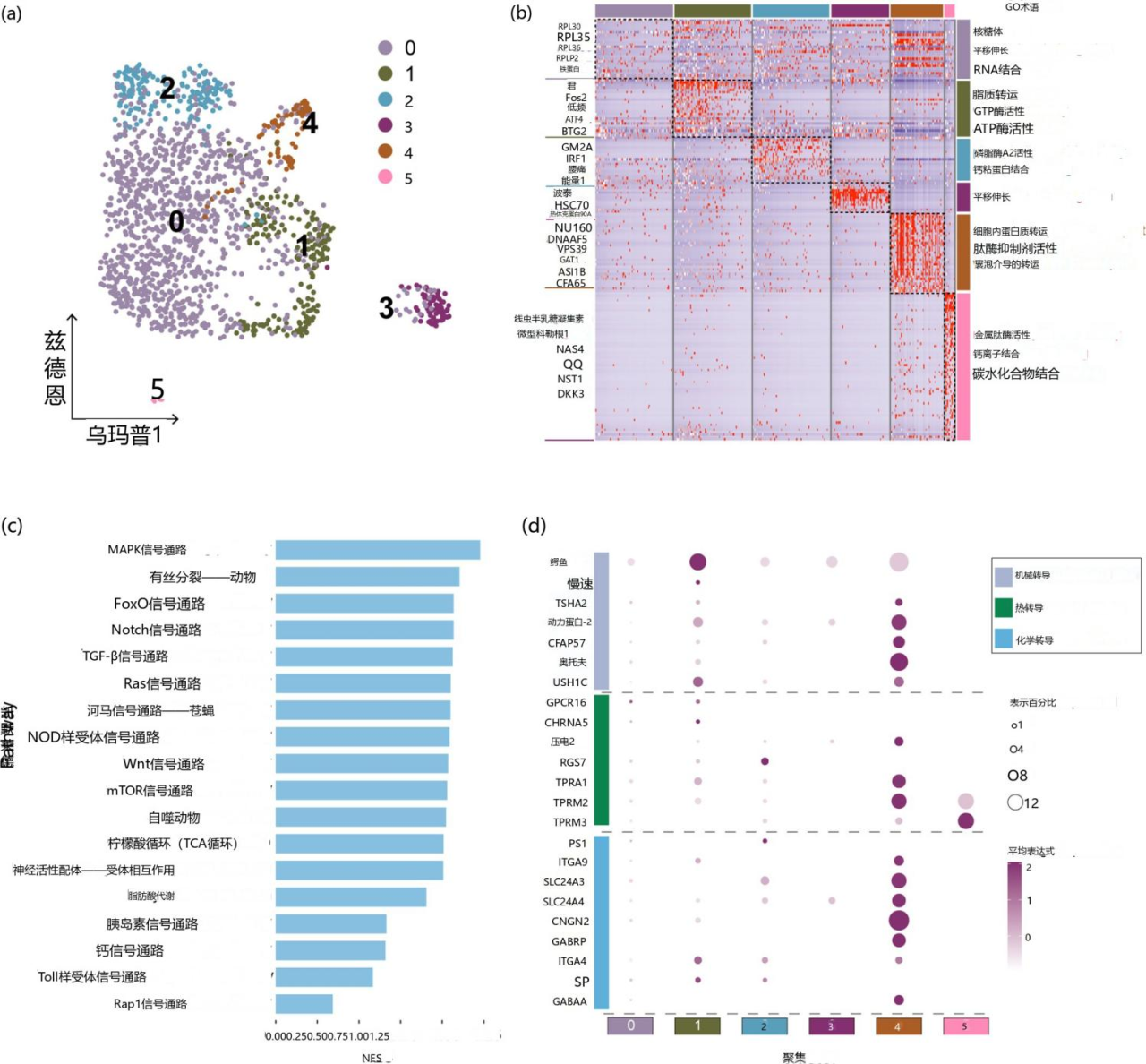
图 4 水螅态A. coerulea细胞图谱
总结
休眠是一种生存和繁殖策略,使生物体能够承受不利的环境条件。本研究证明在Aurelia属中,足囊的形成是由饥饿诱导的,同时较高的温度会加速足囊的形成。在这项研究中,BD Rhapsody单细胞平台发挥了重要作用,快速准确地捕获了上万个单细胞并进行测序,为针对足囊形成和维持阶段的细胞类型特异性转录景观绘制出了第一个高分辨率的单细胞图谱,提高了对调节休眠足囊形成和维持的分子机制的理解,阐明了潜在的调节途径和环境因素之间复杂的相互作用,并为了解不同生物体和生态栖息地的适应性生存策略的深远影响提供了宝贵的见解。
原文链接:https://doi.org/10.1111/mec.17249








 沪公网安备31011502400759号
沪公网安备31011502400759号
 营业执照(三证合一)
营业执照(三证合一)




