







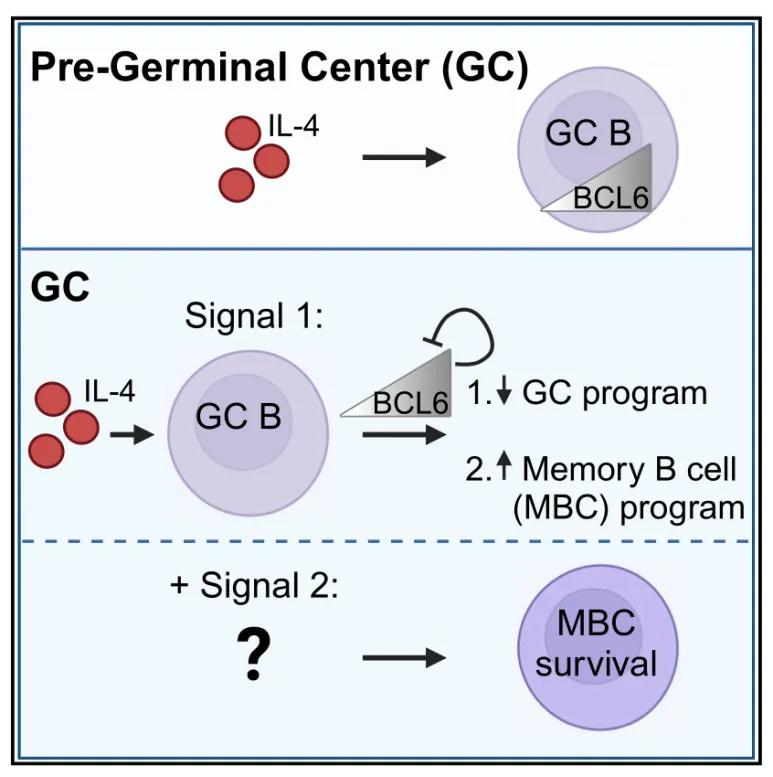
一、抗病毒早期免疫调控:IL-4/BCL6 启动GC形成
抗病毒早期,自然杀伤 T 细胞(NKT)分泌 IL-4,促进 B 细胞表达 BCL6 并诱导GC形成。BCL6 通过抑制相关抑制物转录,将 B 细胞与滤泡辅助 T 细胞(TFH)滞留于 GC 内,阻止其终末分化。研究证实,IL-4 过量会减少记忆 B 细胞(MBC)生成、降低选择严格性,但其潜在机制尚未明确。
二、GC功能与核心谜题:BCL6 下调信号如何启动?
GC是 B 细胞滤泡内高度组织化的结构。B 细胞在此经历体细胞高频突变与类别转换重组,实现 BCR 多样化,经选择后成熟为抗体分泌型浆细胞或长期存活的MBC,通过体液免疫抵御外来抗原侵袭。GC 的启动、维持与溶解,依赖 B 细胞与 TFH、滤泡树突细胞(FDC)等细胞的时空复杂相互作用(部分机制已明确),但 B 细胞如何退出 GC 状态形成 MBC 仍不清晰。目前普遍认为,抑制性转录因子 BCL6 是 GC 维持的关键,其表达下调是 MBC 形成的必要条件,但启动 BCL6 下调的具体信号尚未明确。阐明这一过程,对优化疫苗开发、制定自身免疫疾病治疗策略具有重要意义。
三、《Immunity》研究突破:IL-4 充当 GC 可调开关,调控 MBC 选择
近日,美国华盛顿大学医学院 Marion Pepper 团队在《Immunity》杂志发表题为《Interleukin-4 downregulates transcription factor BCL6 to promote memory B cell selection in germinal centers》的研究。作者发现,GCB 细胞中的 IL-4 信号可通过负向自动调节直接下调转录因子 BCL6,使细胞脱离 GCB 程序并促进 MBC 形成,同时导致部分 GCB 细胞解体或死亡。此外,研究证实,IL-4 信号过强或过弱均会破坏 MBC 选择严格性。由此,IL-4 对 BCL6 表达的调控可作为 GC 内的可调开关,精准调控 MBC 选择与亲和力成熟过程。

四、IL-4/BCL6 信号轴调控生发中心功能与记忆 B 细胞选择的机制解析
1. 感染模型验证:B 细胞对 IL-4 的感知及 IL-4 对 GC 形成的双重影响
- 实验模型与基础发现:采用疟原虫 y-GP66 感染的红细胞(iRBC)感染 KN2 小鼠(IL-4 启动子调控 huCD2 表达),观察到感染急性期产 IL-4 的 T 细胞数量显著增加。
- B 细胞对 IL-4 的感知能力:通过疟原虫血期抗原 MSP1 四聚体检测证实,初始 B 细胞(naïve B 细胞)和 MSP1 特异性GCB 细胞均表达 IL-4 受体链(IL-4R),说明 B 细胞在分化为 GCB 细胞前后均能感知 IL-4 信号。
- IL-4 对 GC 形成的阶段特异性作用:用 iRBC 感染野生型(WT)小鼠,分别静脉注射 IL4C(重组 IL-4 与抗 IL-4 抗体复合物)和 PBS 作为对照。感染第 8 天(GC 形成早期),IL4C 组 MSP1 特异性 GCB 细胞数显著增多,证实 IL-4 可促进早期 GC 形成;感染第 10 天(GC 已形成),IL4C 组 MSP1 特异性 GCB 细胞数明显下降,且记忆 B 细胞(MBC)未同步升高,提示 IL-4 在 GC 形成后可能限制 GC 反应。
2. IL-4 下调 BCL6 调控 GCB 细胞命运,调控依赖 BCL6 基础水平
- IL-4 限制 GC 反应的验证:脾脏成像显示,IL4C 组 GC 体积显著小于 PBS 组,证实 IL-4 可限制 GC 反应。
- IL-4 介导 BCL6 下调及 GCB 细胞分化:分析 MSP1 特异性 B 细胞发现,IL4C 组 GC 转化状态 B 细胞的 BCL6 显著下调;流感模型验证,GCB 细胞及 GC 转化状态 B 细胞的 BCL6 均降低。流式染色显示,IL-4 下调 BCL6 可促使 GCB 细胞退出 GC 状态,与 MBC 标志物 CD73 共表达,向 MBC 方向分化。
- 调控依赖 BCL6 基础水平:初始 B 细胞经 CD40 抗体联合 IL-4 刺激,BCL6 表达显著增高;相同条件下,GCB 细胞的 BCL6 表达几乎消失,说明调控依赖细胞自身 BCL6 基础水平。
- 核心机制:IL-4 激活转录激活因子 STAT6 和 IRS2,触发 BCL6 负向自动调节,使细胞退出 GC 状态,但这类细胞无法存活为 MBC。
3. IL-4 介导 BCL6 下调伴随 BCL2 缺失,促进 GCB 细胞死亡
- BCL2 的表达特征与功能:抗凋亡蛋白 BCL2 在初始 B 细胞中高表达,进入 GC 后下调,GCB 细胞分化为 MBC 时再次上调,且 BCL2 可抑制 BCL6。
- IL-4 调控下的 BCL2 表达:染色结果显示,IL-4 下调 BCL6 时,并未同步上调 BCL2。
- 细胞死亡的诱因与验证:BCL2 缺失导致 GCB 细胞死亡增加;BCL2 转基因小鼠实验进一步证实,该细胞死亡由 IL-4 介导。
4. IL-4 信号强度调控 MBC 选择严格性,过强或过弱均降低抗体亲和力
- MBC 分化的核心前提:GCB 细胞需经体细胞高频突变获得高亲和力 BCR,才能进一步分化为 MBC;正常情况下,GCB 细胞会上调 BCL2 以维持 MBC 长期存活。
- 过量 IL-4 的影响:重复刺激模型检测显示,过量 IL-4 存在时,形成的 MBC 亲和力较低。
- IL-4 信号缺失的影响:感染 WT 小鼠与 IL-4R 敲除(IL-4RKO)小鼠后,IL-4RKO 组感染 60 天的 GC B 细胞数显著增加,但血清抗 MSP1 特异性抗体水平与 WT 组无差异,表明 IL-4 不影响浆细胞选择;二次感染实验显示,IL-4RKO 小鼠血清相关抗体信号显著减少,提示其 MBC 衍生抗体亲和力低于 WT 小鼠。
- 核心结论:IL-4 可严格调控 GC 中 MBC 的选择,信号过强或过弱均会导致选择严格性丧失。
5. 核心模型:IL-4 以 BCL6 表达量为开关,调控 B 细胞进出 GCB 状态
- 调控核心逻辑:IL-4 对 B 细胞进出 GCB 状态的调控,取决于细胞自身 BCL6 的表达量。不表达或低表达 BCL6 的细胞(如初始 B 细胞),响应 IL-4 信号后会增强 BCL6 表达;高水平表达 BCL6 的细胞(如成熟 GCB 细胞),响应 IL-4 信号后会下调 BCL6。
- 具体调控过程:免疫反应早期,自然杀伤 T 细胞(NKT)或滤泡辅助 T 细胞(TFH)产生的 IL-4,作用于初始 B 细胞和早期激活 B 细胞,促进 BCL6 表达,推动其向 GCB 细胞迁移;GC 形成后,TFH 细胞持续产生大量 IL-4,作用于 GCB 细胞,使 BCL6 表达升高至触发负向自动调节的阈值,进而下调 BCL6,释放已获得足够辅助信号的 GCB 细胞,促进高亲和力 MBC 形成。
- 关键调控节点:BCL6 作为响应 IL-4 信号的可调开关,是 GC 中 MBC 选择的关键调控节点。
乐备实多款Panel助力免疫研究,欢迎咨询~








 沪公网安备31011502400759号
沪公网安备31011502400759号
 营业执照(三证合一)
营业执照(三证合一)




