







一、引言
EGF于上世纪被Stanley Cohen与Rita Levi-Montalcini发现并获诺贝尔生理学奖,是EGF生长因子家族原型分子,同家族包含TGF-α、HB-EGF等功能性同源蛋白。自被发现以来,EGF从基础分子生物学研究逐步延伸至创面修复、肿瘤药理、消化内科等多领域转化应用。生理条件下,EGF精准调控上皮细胞更新、皮肤及黏膜组织修复、乳腺与骨骼发育;而EGF/EGFR通路持续异常活化是上皮源性肿瘤恶性增殖、侵袭转移的核心驱动因素,同时参与慢性炎症、器官纤维化的病理演变。随着重组蛋白表达技术(HEK293真核表达系统)成熟,商品化重组人源EGF细胞因子试剂已广泛用于体外细胞药理学试验,重组EGF外用制剂也实现临床落地。本文围绕EGF基础理化、受体作用、信号调控、疾病关联及临床转化五大维度展开系统性综述。
二、EGF基础理化性质与体内合成分泌特征
(一)分子结构与理化属性
成熟EGF为含53个氨基酸残基的单链耐热多肽,分子量约6 kDa;分子内形成3对特征性分子内二硫键,构筑保守EGF样结构域,该结构是EGF维持生物活性、特异性结合EGFR受体的必需结构基础。EGF家族全部成员均依托该保守结构域实现受体识别,二硫键断裂将直接导致EGF完全丧失生物学活性。
(二)前体合成与组织来源
EGF在体内以跨膜前体Pro-EGF形式合成,前体多肽全长含1217个氨基酸残基,主要在哺乳动物颌下腺、肾脏、十二指肠布氏腺(Brunner腺)组织细胞中表达。Pro-EGF锚定细胞膜后,经组织特异性蛋白酶定点酶切水解,胞外片段脱落形成可溶性成熟EGF;区别于多数瞬时分泌型细胞因子,机体合成的大量EGF储存于组织细胞胞浆颗粒中,受生理信号调控后阶段性释放,非持续性胞外分泌。
三、EGF-EGFR受体结构及配体-受体活化机制
(一)EGFR(ErbB1/HER1)分子分区结构
EGFR隶属于ErbB酪氨酸激酶受体(RTK)超家族,单体受体分为三段结构:
(1)胞外区:由I~IV四个独立结构域组成,无配体结合时,结构域II与IV相互贴合形成自抑制拴系(tethered)构象,阻断受体二聚化;I、III结构域为EGF特异性结合位点。
(2)跨膜区:单次跨膜α螺旋结构,串联胞外配体结合区与胞内激酶功能区。 3. 胞内区:保守酪氨酸激酶催化结构域+C端多酪氨酸残基尾段,C端酪氨酸磷酸化后形成下游信号蛋白锚定结合位点。
(二)EGF诱导受体二聚化活化过程
成熟EGF结合EGFR胞外I、III结构域,诱发受体构象重排,解除胞外区自抑制状态,暴露结构域II二聚化臂;游离受体单体随即发生同源二聚(EGFR-EGFR),或与ErbB2(HER2)等家族成员形成异源二聚体。受体二聚化是胞内激酶激活的先决条件,二聚体形成后两单体胞内激酶区相互靠近,发生交叉自体酪氨酸磷酸化,C端多个酪氨酸位点被磷酸修饰,形成SH2/PTB结构域蛋白的招募锚定位点,正式启动下游信号级联反应。
四、EGF/EGFR下游四大核心信号传导通路
磷酸化EGFR通过锚定适配蛋白启动四条经典胞内信号通路,各通路分工调控细胞不同生理功能:
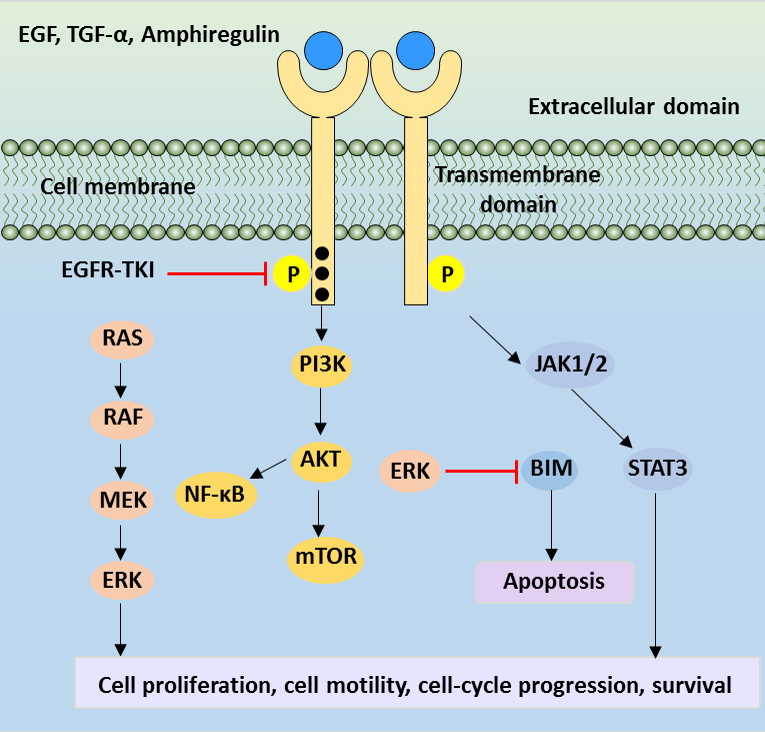
EGF/EGFR信号传导通路
(一)RAS-MAPK通路(增殖分化主控通路)
磷酸化EGFR招募Grb2-SOS复合物,SOS作为鸟苷酸交换因子激活小G蛋白Ras;活化Ras逐级激活Raf→MEK→ERK激酶级联,活化ERK入核调控靶基因转录,主导细胞增殖、周期调控与细胞分化,该通路异常持续激活是肿瘤无限增殖关键诱因。
(二)PI3K-Akt通路(细胞存活抗凋亡通路)
EGFR直接或通过Gab1适配蛋白招募磷脂酰肌醇3激酶(PI3K),PI3K催化细胞膜PIP2转化为PIP3;PIP3募集并活化Akt激酶,活化Akt通过抑制促凋亡蛋白、上调抗凋亡基因,抑制细胞凋亡、提升细胞存活能力,肿瘤中该通路过度活化助力癌细胞逃逸凋亡。
(三)JAK-STAT通路(基因表达调控通路)
在上皮、造血等特定细胞亚型中,活化EGFR间接激活JAK激酶,进一步磷酸化STAT转录因子;磷酸化STAT形成同源二聚体入核,靶向结合靶基因启动子,调控炎症、增殖相关基因表达,参与组织稳态与炎症反应调控。
(四)PLC-γ通路(细胞运动与形态调控通路)
PLC-γ被磷酸化EGFR招募活化,水解细胞膜PIP2生成IP3与DAG:IP3促使内质网钙离子外流升高胞内钙浓度;DAG激活蛋白激酶PKC,二者协同调控细胞骨架重塑、细胞迁移与形态改变,参与创面修复细胞游走及肿瘤侵袭转移。
(五)EGF信号负反馈终止机制
机体通过多重负调控精准限制EGF信号强度,避免通路持续异常激活:
① 配体-受体复合物经网格蛋白介导胞吞入胞;
② EGFR泛素化标记后被溶酶体/蛋白酶体降解;
③ 蛋白酪氨酸磷酸酶(PTPs)催化受体C端酪氨酸去磷酸化,终止下游信号招募。
上述任一调控环节缺失,均可诱发EGFR持续活化致病。
五、EGF/EGFR信号异常与相关疾病发生机制
EGF通路具有生理修复、病理促病变的双向作用,信号稳态失衡介导四类重大疾病发生发展。
(一)恶性上皮源性肿瘤
非小细胞肺癌、结直肠癌、胶质母细胞瘤、头颈部鳞癌等上皮来源肿瘤中,EGFR基因扩增、体细胞激活突变、蛋白过表达频发,造成EGFR无需EGF配体即可自主二聚活化,下游RAS-MAPK、PI3K-Akt通路永久激活,驱动肿瘤细胞无限增殖、抗凋亡、侵袭转移及肿瘤新生血管生成。
临床靶向治疗:
① 大分子单克隆抗体:西妥昔单抗、帕尼单抗,靶向胞外EGFR阻断EGF结合;
② 小分子酪氨酸激酶抑制剂(TKI):吉非替尼、厄洛替尼、奥希替尼,胞内竞争性抑制EGFR激酶活性,是晚期EGFR突变肿瘤一线靶向用药。
(二)急慢性皮肤创面损伤
生理浓度EGF可靶向促进角质形成细胞、成纤维细胞、血管内皮细胞增殖与定向迁移,加速表皮爬行再生、新生血管形成与肉芽组织构建。重组人表皮生长因子(rhEGF)已获批外用制剂,临床用于烧伤创面、糖尿病难治性足溃疡的局部修复,显著缩短创面愈合周期,是再生医学成熟落地产品。
(三)炎症性肠病(IBD)
溃疡性结肠炎等炎症性肠病以肠道上皮屏障破损、黏膜糜烂溃疡为核心病理特征;EGF通过加速小肠、结肠上皮细胞增殖再生,修复受损肠黏膜屏障,减少肠道菌群异位诱发的持续炎症。临床基础试验证实外源性补充EGF可促进肠道黏膜愈合,为炎症性肠病辅助治疗提供新方向。
(四)慢性器官纤维化疾病
短期生理性EGF利于组织损伤修复,但长期过度激活EGFR信号持续刺激成纤维细胞异常增殖,促使胶原、纤连蛋白等细胞外基质过量沉积,诱发肺间质纤维化、肝纤维化等难治性纤维化病变,是慢性脏器损伤向纤维化进展的重要分子诱因。
六、研究总结与未来展望
(一)现有研究总结
EGF作为经典生长因子,其分子结构、受体互作、四条核心信号通路的基础机制已被系统阐明;生理层面是机体上皮组织稳态维持、损伤修复必需因子,病理层面通路异常是肿瘤、纤维化、慢性炎症的关键致病靶点。依托HEK293真核表达平台的重组人源EGF商品化试剂成熟,满足体外细胞试验需求;rhEGF外用修复制剂、EGFR靶向抗肿瘤药实现产业化临床应用,打通基础研究向临床转化路径。但仍存在短板:
① 靶向药物耐药突变频发(如EGFR T790M、C797S突变)限制抗肿瘤药效;
② 全身给药EGF半衰期短、易被酶解,难以实现内脏黏膜靶向给药;
③ 纤维化中EGFR精准抑制药物仍缺乏临床转化产品。
(二)未来研究方向
(1)抗肿瘤药物研发:开发新一代EGFR不可逆抑制剂、EGFR/HER2双特异性抗体、ADC偶联药物,攻克靶向药获得性耐药;推进靶向联合疗法(靶向药+免疫检查点抑制剂/放化疗)提升实体瘤客观缓解率。
(2)再生医学制剂优化:依托纳米载体、生物水凝胶、缓释微球技术改造EGF剂型,延长体内半衰期,实现肠道、角膜等深部组织靶向缓释给药,拓展炎症性肠病、角膜溃疡等适应症。
(3)抗纤维化靶向新药:筛选选择性EGFR小分子抑制剂,特异性阻断成纤维细胞异常活化,研发肺、肝纤维化靶向候选药物。
(4)基础机制深挖:探索非经典EGFR激酶非依赖信号通路、EGF与肠道菌群互作新机制,挖掘全新疾病干预靶点。
七、EGF 相关检测服务哪里有?
乐备实生物 LabEx 可提供专业的 EGF/EGFR 通路相关科研检测服务。平台搭载 Luminex MSD、抗体芯片、WB 蛋白定量、免疫组化等成熟技术,可开展 EGF 家族因子定量、EGFR 表达与磷酸化检测、通路关键蛋白筛选、多标志物联合测定等实验,全面适配 EGF 通路机理探究、药效评估、疾病标志物筛选等各类科研实验需求。








 沪公网安备31011502400759号
沪公网安备31011502400759号
 营业执照(三证合一)
营业执照(三证合一)




