







一、引言
自噬源自希腊语“自我吞噬”,是细胞进化保留的自我修复与应激生存策略,兼具胞内垃圾清除、营养循环、抗感染、抗衰老多重生理功能。1963年Christian de Duve首次定名自噬,后续数十年围绕自噬分型、分子通路的基础研究逐步完善,2016年大隅良典阐明自噬核心分子机制推动该领域跨越式发展。传统研究多聚焦自噬基础生化过程,2026年我国中科院、北大、南农多个课题组接连在顶级期刊发表原创成果,从内质网自噬启动信号、自噬体-溶酶体融合调控、代谢-自噬轴抗病毒三大维度突破既往研究瓶颈,同时自噬靶向小分子药物与仿生纳米制剂陆续进入临床前验证阶段,推动自噬研究由基础机理探索迈向精准疾病干预。本文基于现有研究成果,系统综述自噬分类、分子机制、最新科研突破与临床转化研究。
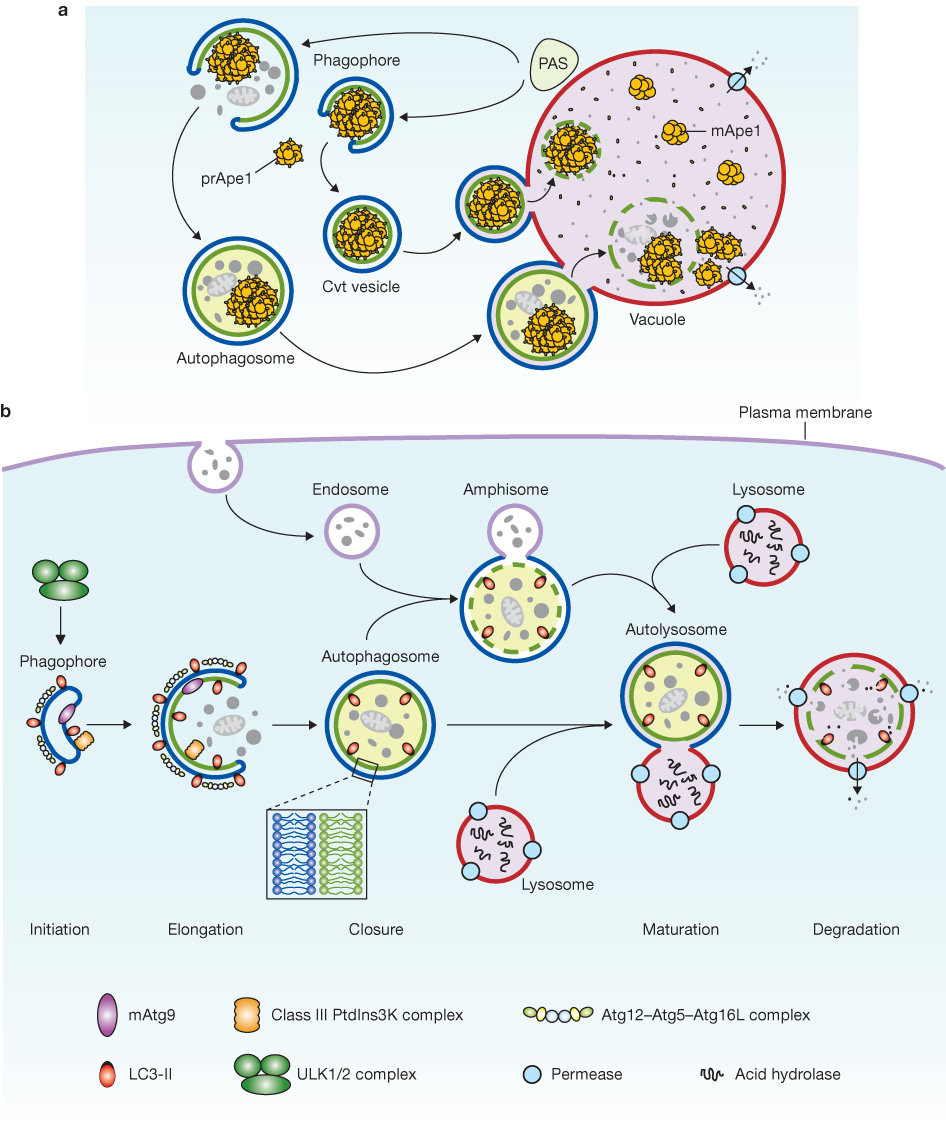
细胞自噬基本过程
二、自噬的定义与分类
(一)自噬基本定义
自噬是细胞在营养匮乏、能量短缺、氧化应激、病原体入侵等环境刺激下,以双层膜结构包裹胞内异常组分形成自噬体,自噬体与溶酶体融合生成自噬溶酶体,依靠酸性水解酶降解内容物,释放氨基酸、脂肪酸等小分子物质供细胞重新利用的保守生理过程,是维持细胞稳态、抵御病理损伤的核心调控通路。
(二)自噬四大分型
依据底物转运入溶酶体的路径与底物选择性,自噬分为巨自噬、微自噬、分子伴侣介导自噬、选择性自噬四类:
(1)巨自噬(Macroautophagy):经典研究最充分的自噬亚型,依靠隔离膜包裹胞质组分形成双层膜自噬体,再与溶酶体融合降解底物,常规语境下“自噬”多指巨自噬。
(2)微自噬(Microautophagy):无需生成自噬体,溶酶体膜直接向内凹陷或向外凸起,包裹细胞质底物进入溶酶体腔完成降解。
(3)分子伴侣介导的自噬(CMA):高度选择性蛋白降解途径,分子伴侣Hsc70特异性识别携带KFERQ保守基序的靶蛋白,介导底物结合溶酶体膜LAMP-2A受体并跨膜入溶酶体降解。
(4)选择性自噬(Selective Autophagy):靶向特定细胞器或外源病原体的精准自噬,细分线粒体自噬(清除损伤线粒体)、内质网自噬(降解异常内质网)、过氧化物酶体自噬、核糖体自噬、异源自噬(清除胞内细菌、病毒),是细胞靶向清除病理性组分的关键形式。
三、自噬核心分子调控机制
自噬全过程由30余种ATG(自噬相关)蛋白分级协同调控,整体划分为起始、成核、膜延伸、成熟降解四个关键阶段,mTOR、AMPK是上游核心信号开关。
(一)起始阶段:ULK1复合物活化(自噬启动开关)
ULK1复合物(ULK1/2+ATG13+FIP200+ATG101)决定自噬是否启动。营养充足时,mTORC1激酶磷酸化ULK1、ATG13,持续抑制自噬起始;饥饿、能量匮乏条件下,mTORC1被抑制、活性下调,ULK1去磷酸化活化,依次磷酸化下游ATG13与FIP200,开启自噬程序。同时,能量不足激活AMPK,通过磷酸化ULK1的Ser467、Thr574位点直接激活ULK1,协同诱导自噬。
(二)成核阶段:PI3KC3复合物组装
活化的ULK1招募III型磷脂酰肌醇3激酶(PI3KC3)复合物(VPS34、Beclin1、ATG14L);VPS34催化生成PI3P,富集PI3P的膜位点招募DFCP1、WIPI家族蛋白,在内质网-线粒体接触位点组装前自噬体结构(PAS),搭建隔离膜合成平台。AMPK可直接磷酸化Beclin1(Ser91/94)与VPS34,增强PI3KC3复合物促自噬活性。
(三)延伸阶段:两套泛素样连接系统介导隔离膜延展
隔离膜双向延伸依赖两套保守泛素化修饰通路:
(1)ATG12-ATG5-ATG16L1系统:ATG7(E1酶)、ATG10(E2酶)催化ATG12与ATG5共价结合,复合物进一步结合ATG16L1并锚定隔离膜外膜,促进膜结构扩张。
(2)LC3脂化修饰通路:胞质游离LC3-I经ATG4剪切,在ATG7、ATG3催化下与磷脂酰乙醇胺(PE)偶联形成膜结合型LC3-II;LC3-II定位于自噬体双层膜,既是自噬体经典标志物,也参与膜延伸与底物识别。
(四)成熟与降解阶段:自噬体-溶酶体融合与底物水解
成熟自噬体依靠细胞微管系统转运至溶酶体,在SNARE复合体、Rab GTP酶协同作用下完成膜融合形成自噬溶酶体;溶酶体内组织蛋白酶等酸性水解酶降解囊内底物,代谢小分子被转运至胞质循环利用,完成一轮自噬流转。
四、2026年自噬领域三项关键性原创研究进展
2026年4月,国内三支科研团队分别在《Molecular Cell》《Nature Communications》《Autophagy》刊发突破性成果,从内质网自噬启动、自噬体融合转运、代谢-自噬抗病毒三个方向补齐领域关键空白。
(一)中科院张宏团队揭示Ca²⁺与机械通道调控内质网自噬启动机制(Molecular Cell,2026)
内质网是细胞主要钙库,内质网自噬是清除损伤内质网、维持钙稳态的关键选择性自噬,但其启动信号与自噬体膜来源长期不明。张宏团队利用超分辨活细胞多模态成像证实:饥饿、胆固醇紊乱、钙超载应激下,内质网高钙片状亚域发生结构重塑,自噬体膜直接来源于内质网膜重构,排除细胞膜来源假说;机械敏感通道PIEZO1、TRPV1富集于高钙内质网膜,感知钙浓度波动触发局部瞬时钙释放,钙瞬变诱导FIP200复合物发生液-液相分离,进而启动内质网自噬。该研究明确内质网自噬上游钙信号通路,为钙稳态失衡诱发的神经退行性疾病、恶性肿瘤提供全新药物靶点。
(二)北大陈建国课题组阐明STX17乙酰化修饰调控自噬体-溶酶体融合转运机制(Nat Commun,2026.4.30)
SNARE蛋白STX17是自噬体与溶酶体融合的核心分子,但静息态STX17弥散分布于内质网、线粒体与胞质,自噬激活时靶向转运机制此前未被阐明。研究借助CRISPR定点敲入、单分子荧光追踪技术证实:介导膜融合的功能性STX17主要储备于胞质可溶性库,非细胞器膜来源;饥饿诱导乙酰转移酶GCN5催化STX17第254位赖氨酸(K254)乙酰化,去乙酰化酶SIRT1可逆调控该修饰;K254乙酰化显著提升STX17与肌动蛋白马达蛋白MYO6结合能力,由MYO6介导胞质STX17定向转运至自噬体膜。该乙酰化动态开关完善自噬晚期调控网络,助力神经退行病、肿瘤的自噬靶向干预研发。
(三)南农周斌团队阐明DHODH介导代谢-自噬轴调控瘟病毒复制(Autophagy,2026.4)
二氢乳清酸脱氢酶(DHODH)是嘧啶从头合成限速酶,该研究首次建立代谢-选择性线粒体自噬-病毒复制关联通路:猪瘟病毒(瘟病毒属)NS4A蛋白结合DHODH并将其招募至线粒体,NS4A的LIR基序激活线粒体自噬,病毒借助活化自噬流获取嘧啶原料完成基因组复制;该调控模式存在病毒属特异性:瘟病毒依赖DHODH维持自噬流,黄病毒则利用自噬阻断外源嘧啶合成。补充外源性嘧啶前体可逆转异常线粒体自噬、恢复自噬体-溶酶体融合效率,为抗瘟病毒新药开发开辟代谢-自噬靶向新策略。
五、总结与研究展望
自噬作为细胞与生俱来的自我救赎机制,从基础生化过程逐步延伸至衰老、肿瘤、神经病变、病原感染多类重大疾病的病理调控靶点。2026年国内三项标志性成果打通内质网自噬启动、自噬体膜融合、代谢-自噬抗病毒三大关键环节,完善自噬上下游调控网络;同时多款精准靶向自噬候选药物加速落地,标志自噬转化医学进入高速发展期。
六、细胞自噬研究实验代测服务哪里有?
自噬相关机制解析、衰老标志物筛选与候选药物药效评价高度依赖高通量、多维度的蛋白检测技术支撑,乐备实生物(LabEx)依托成熟的 Luminex 多因子芯片、流式多因子、抗体芯片、蛋白定量及翻译后修饰组学平台,可一站式承接衰老与自噬方向科研服务,覆盖自噬通路关键蛋白定量、细胞因子谱检测、靶蛋白修饰分析、疾病标志物筛选等实验内容,能够适配自噬机理探究、通路验证、抗衰老药物体内外功效评估等全链条科研需求,为相关基础课题落地与靶向药物研发提供可靠的实验技术保障,助力自噬科研成果从理论研究稳步走向转化应用。








 沪公网安备31011502400759号
沪公网安备31011502400759号
 营业执照(三证合一)
营业执照(三证合一)




