







引言
二代测序技术的突破,让人类基因组中曾被忽视的 “暗物质”—— 非编码 RNA(ncRNA)逐渐显露重要作用。研究发现,人类基因组仅约 10% 转录本编码蛋白质,其余 90% 均为非编码 RNA,其中小非编码 RNA(sncRNA,长度≤400nt) 凭借精准、多样的调控能力,成为基因表达调控网络的核心参与者。这类 RNA 不编码蛋白质,却能在转录、转录后及翻译水平调控基因表达,参与细胞增殖、分化、凋亡乃至肿瘤发生等关键过程。目前,miRNA(微小 RNA)是研究最成熟的 sncRNA,而 tiRNA/tRF(tRNA 衍生片段)作为新兴成员,其调控潜力正逐步被挖掘。小非编码RNA的定义与分类,小非编码RNA是一类不编码蛋白质、但具有重要调控功能的短RNA分子,通常小于200个核苷酸。其主要类型包括miRNA(微RNA)、siRNA(小干扰RNA) 和piRNA(Piwi互作RNA) 等,它们虽然在生成和作用机制上有所不同,但共同构成了基因表达调控网络的核心组成部分。本文将聚焦这两类典型 sncRNA,解析其分子特征、生物发生路径及基因调控机制,揭示它们在生命活动中的关键作用。
一、miRNA:基因表达的经典 “转录后刹车”
自 1993 年 Ambros 团队在线虫中发现首个 miRNA(lin-4)以来,这类小分子 RNA 的调控机制已被深入研究。人类基因组中已注释的 miRNA 超 2000 种,每种可靶向数十至数百个 mRNA,构建起复杂的调控网络。
1.1 miRNA 的分子特征与生物发生
miRNA 是长度 18-24nt 的单链 RNA,核心特征是 5’端第 2-8nt 的 “种子序列”—— 这是其识别靶 mRNA 的关键。其生物发生需经 “核内加工 - 胞质成熟” 两步:首先,miRNA 基因转录生成含茎环结构的初级 miRNA(pri-miRNA),在核内被 Drosha 酶与辅助因子 Pasha 切割,生成 60-110nt 的前体 miRNA(pre-miRNA);随后,pre-miRNA 经 Exportin 5 转运至细胞质,被 Dicer 酶切割颈环结构,形成 22nt 左右的双链 miRNA(miRNA/miRNA*);最终,双链进入 RNA 诱导的沉默复合物(RISC),互补链(后随链)被 AGO2 蛋白降解,向导链(成熟 miRNA)保留,完成成熟。值得注意的是,向导链的选择由双链 5’端稳定性决定:RISC 更倾向选择 5’端 AT 含量高、碱基配对弱的链,这一机制确保了调控特异性,避免非特异性干扰。
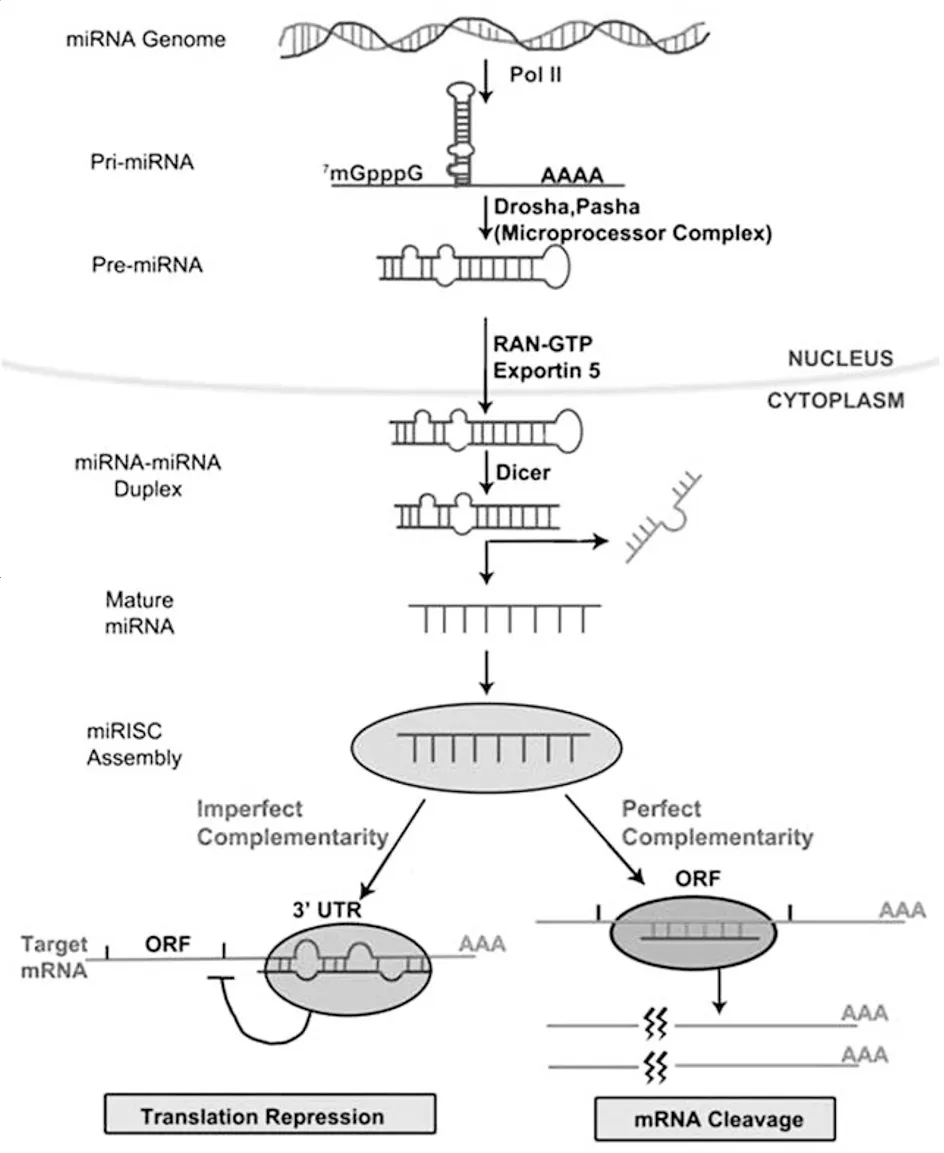
图1. miRNA合成过程
1.2 miRNA 的基因调控机制
miRNA 通过 RISC 与靶 mRNA 结合,在转录后水平抑制基因表达,具体依赖 “种子序列” 与靶 mRNA 的互补程度,主要分为三条通路:一是抑制翻译起始,当 miRNA 与靶 mRNA 3’非翻译区(3’UTR)不完全互补时,AGO2 蛋白竞争结合 mRNA 的 5’帽子结构,阻止核糖体组装;二是抑制核糖体延伸,RISC 结合后通过变构效应阻止核糖体在 mRNA 上移动,导致未完成的多肽链降解;三是降低 mRNA 稳定性,RISC 招募去帽酶与脱腺苷酸酶,去除 mRNA 的 5’帽子与 3’多聚腺苷酸尾,加速其降解。此外,若 miRNA 与靶 mRNA 完全互补,AGO2 可直接切割 mRNA,引发快速降解。
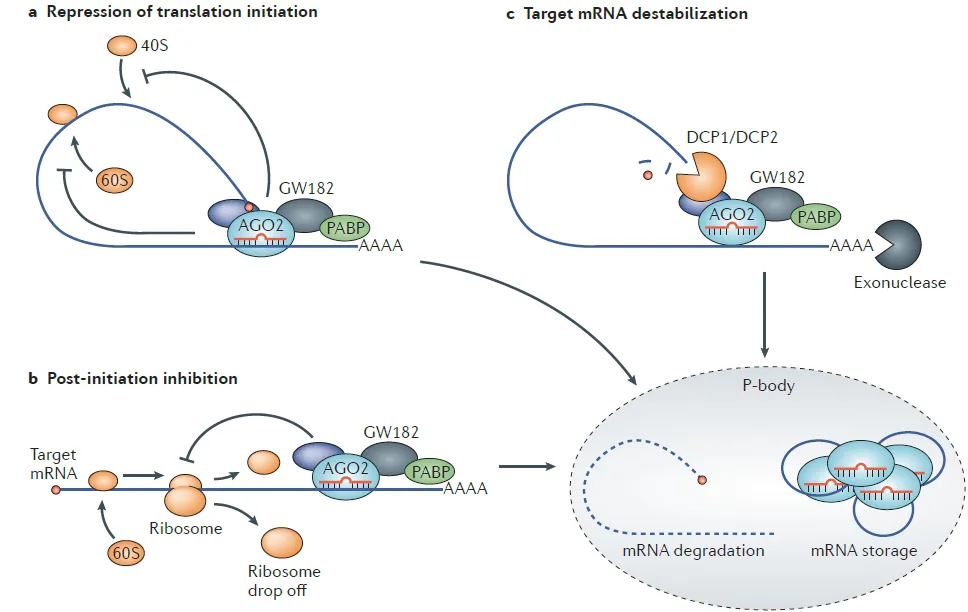
图2. miRNA对mRNA的调节过程
二、tiRNA/tRF:源于 tRNA 的新型 “调控新秀”
除 miRNA 外,tRNA 衍生片段(tiRNA/tRF)是近年发现的新型 sncRNA。tRNA 本是蛋白质合成中转运氨基酸的分子,而 tiRNA/tRF 是 tRNA 在应激条件或特定酶切下产生的片段,其调控模式兼具 miRNA 的相似性与独特性。
2.1 tiRNA/tRF 的分子特征与分类
成熟 tRNA 长度约 79nt,呈 “三叶草” 二级结构,人类基因组中约 506 个基因编码 49 种 tRNA。tiRNA/tRF 按切割位置与大小可分为两类:一是 tiRNA(tRNA 半分子),由血管生成素在 tRNA 反密码子环切割产生,长度 29-50nt,分 5’tiRNA(5’端至反密码子环)与 3’tiRNA(反密码子环至 3’端),5’端为羟基,成熟不依赖 Dicer;二是 tRF(tRNA 衍生片段),长度 14-30nt,5’端含磷酸,分 tRF-5(源于 tRNA 5’端)、tRF-3(源于 3’端)、tRF-2(仅含反密码子区)与 tRF-1(源于前体 tRNA 3’端拖尾序列)。
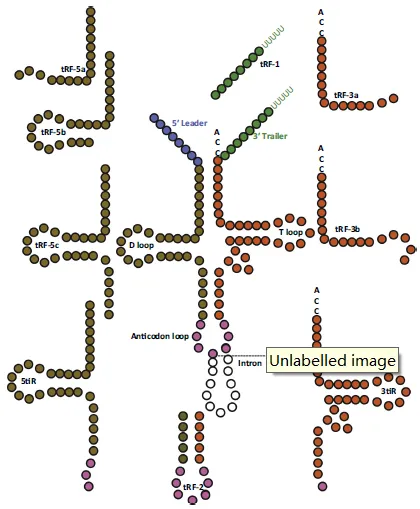
图3. tRF的分类
2.2 tiRNA/tRF 的生物发生与调控功能
tiRNA/tRF 的生物发生仍有较多未知,但核心特征已明确:多数不依赖 Dicer,由 angiogenin 或其他 RNA 酶介导成熟;成熟后可与 AGO1、AGO3、AGO4 等蛋白结合,形成类 RISC 复合物,通过 “种子序列” 结合靶 mRNA,类似 miRNA 的调控模式。
更重要的是,tiRNA/tRF 的调控功能更广泛:除转录后抑制外,还可在转录水平结合基因启动子抑制转录,在表观遗传水平调控组蛋白修饰(如 H3K9me3、H3K27ac);同时参与细胞周期调控(如抑制 RPA1 导致 G1 期阻滞)、肿瘤发生(促进或抑制癌细胞增殖)及应激响应(抑制渗透压诱导的凋亡),是基因调控网络中兼具多样性的新型因子。
三、总结与展望
小非编码 RNA(miRNA、tiRNA/tRF 等)的研究颠覆了 “蛋白质中心论”,证实它们是基因表达的 “精细调节器”。目前,miRNA 的机制已成熟,其作为疾病标志物与治疗靶点的应用进入临床试验;而 tiRNA/tRF 的生物发生、靶标识别仍需深入探索。
未来,随着单细胞测序、空间转录组等技术的发展,更多新型 sncRNA 可能被发现 —— 例如,基因组中异染色质区域或存在低水平转录,其异常中止的转录本可能加工为新型 sncRNA,参与表观遗传调控。这些发现将进一步完善基因调控网络认知,为疾病机制研究与治疗提供新方向。
小非编码RNA的临床应用前景,基于其精准的基因调控能力,小非编码RNA已成为极具潜力的新型治疗靶点和生物标志物。例如,人工合成的miRNA类似物或抑制剂(antagomir)正在被开发为新一代 therapeutics,用于治疗那些由特定基因异常表达所导致的疾病。
乐备实是国内专注于提供高质量蛋白检测以及组学分析服务的实验服务专家,自2018年成立以来,乐备实不断寻求突破,公司的服务技术平台已扩展到单细胞测序、空间多组学、流式检测、超敏电化学发光、Luminex多因子检测、抗体芯片、PCR Array、ELISA、Elispot、PLA蛋白互作、多色免疫组化、DSP空间多组学等30多个,建立起了一套涵盖基因、蛋白、细胞以及组织水平实验的完整检测体系。
我们可提供从样本运输、储存管理、样本制备、样本检测到检测数据分析的全流程服务。凭借严格的实验室管理流程、标准化实验室操作、原始数据储存体系以及实验项目管理系统,已经为超过3000家客户单位提供服务,年检测样本超过100万,受到了广大客户的信任与支持。







 沪公网安备31011502400759号
沪公网安备31011502400759号
 营业执照(三证合一)
营业执照(三证合一)




