







一、引言
基因表达调控是细胞维持生理功能、决定发育命运的核心机制,而 DNA 修饰作为表观遗传领域的关键调控方式,虽不改变 DNA 碱基序列,却能通过化学修饰 “标记” DNA,精准调控基因转录活性。从原核生物的自我防御到真核生物的细胞分化、疾病发生,DNA 修饰始终扮演着 “隐形调控者” 的角色。近年来,随着检测技术的突破,以 N6 - 甲基腺嘌呤(6mA)为代表的新型 DNA 修饰功能被逐步揭示,为理解基因表达调控的复杂性提供了全新视角。
二、DNA 修饰的核心类型与分布特征
2.1 主要 DNA 修饰类型
DNA 修饰类型多样且功能各有侧重。5 - 甲基胞嘧啶(5mC)是哺乳动物中最经典的修饰,由 DNA 甲基转移酶催化形成,广泛参与基因沉默、基因组稳定性维持,是胚胎发育、细胞分化过程中的关键调控因子。N6 - 甲基腺嘌呤(6mA)最早发现于细菌,近年在真核生物中被重新关注,尽管其在真核生物中的含量远低于原核生物,但在基因表达调控中展现出独特作用。此外,由 TET 蛋白家族催化 5mC 生成的氧化修饰(如 5hmC、5fC、5caC),多存在于胚胎干细胞、神经细胞中,与基因激活、DNA 去甲基化过程密切相关。组蛋白修饰关联的 DNA 修饰对基因表达的影响:组蛋白的甲基化、乙酰化等修饰(如 H3K4me3 促激活、H3K9me3 促沉默)会改变染色质结构,与 DNA 甲基化协同作用:开放染色质利于 DNA 与转录蛋白结合,促进基因表达;紧缩染色质则抑制表达。
2.2 基因组分布的区域特异性
DNA 修饰在基因组上的分布并非随机,而是呈现明确的区域偏好。5mC 常富集于基因启动子区的 CpG 岛,该区域的高甲基化通常会抑制转录因子结合,进而导致基因沉默。6mA 在真核生物中多分布于转录起始位点附近及外显子区域,其分布模式与基因表达活性呈正相关。氧化修饰则倾向于集中在活跃转录的基因体区域,可能参与转录过程的动态调控。这种区域特异性分布,是 DNA 修饰能够精准调控基因表达的结构基础。
三、DNA 修饰调控基因表达的关键机制
3.1 直接阻断转录因子结合
DNA 修饰可通过改变 DNA 局部结构,直接影响转录因子与结合位点的相互作用。例如,5mC 修饰会在 CpG 位点引入甲基基团,产生空间位阻,阻碍 AP-2、E2F 等对甲基化敏感的转录因子结合 DNA,进而抑制转录起始复合物的组装,最终导致基因沉默。这种 “物理阻断” 机制,是 DNA 修饰调控基因表达最直接的方式之一。
3.2 重塑染色质结构间接调控
染色质的开放程度决定了转录机器对 DNA 的可及性,而 DNA 修饰可通过影响染色质结构间接调控基因表达。以 6mA 为例,最新研究通过体外核小体组装实验发现,核小体更倾向于缠绕在无 6mA 修饰的 DNA 区域;分子动力学模拟进一步证实,6mA 会改变 DNA 柔性,影响 DNA 与组蛋白的相互作用,从而指导核小体精确定位。体内实验也验证,6mA 缺失会导致核小体排布紊乱,破坏转录组稳定性 —— 核小体定位异常会使染色质局部区域过度紧缩或开放,进而干扰转录因子结合与 RNA 聚合酶招募,影响基因转录效率。此外,5mC 可通过招募 MeCP2 等甲基化结合蛋白,进一步招募组蛋白去乙酰化酶(HDAC),促使组蛋白去乙酰化,增强染色质紧缩程度,间接抑制基因表达,形成 “修饰 - 蛋白招募 - 染色质重塑” 的完整调控链条。
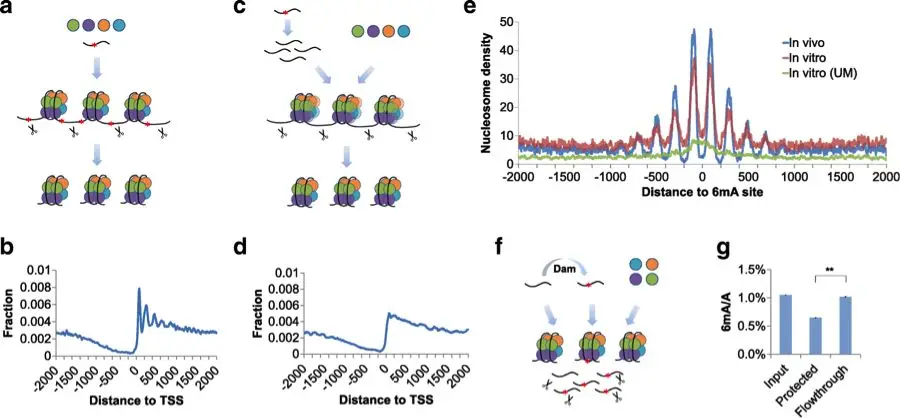
体外核小体组装实验
四、DNA 修饰研究的物种差异
原核生物中,DNA 修饰研究起步早且机制明确,以 6mA 为核心的限制修饰系统是细菌抵御外源核酸入侵的 “防御武器”—— 细菌通过甲基化自身 DNA 标记 “自我”,同时利用限制酶切割未甲基化的外源 DNA,实现对自身基因组的保护。而真核生物中,DNA 修饰研究曾因修饰含量低(如 6mA 占比仅 0.01%-0.1%)、检测技术匮乏而进展缓慢,早期仅在少数物种中发现零星报道,对其具体功能几乎一无所知。真核生物 N6 - 甲基腺嘌呤(6mA)对基因转录组稳定性的影响:真核生物中低含量的 6mA 通过影响 DNA 柔性调控核小体定位,6mA 缺失会导致核小体排布紊乱,干扰转录因子结合与 RNA 聚合酶招募,进而破坏转录组稳定性,影响基因正常转录。
五、总结与展望
DNA 修饰作为基因表达调控的 “隐形之手”,通过直接阻断结合、间接重塑染色质等机制,在生命过程中发挥着不可替代的关键作用。从原核生物的自我防御到真核生物的发育调控,DNA 修饰的功能不断被拓展,但在其调控网络的复杂性方面,仍有诸多问题有待深入探索 —— 例如不同修饰类型间的协同作用机制、特定修饰在疾病发生中的动态变化规律等,仍是未来研究的核心方向。
随着单碱基分辨率修饰检测技术、单细胞表观遗传测序技术的持续发展,我们将能更精准地解析 DNA 修饰的动态变化规律,这不仅为理解肿瘤等疾病中 DNA 修饰异常的发生机制提供支撑,也为开发靶向表观遗传的治疗策略奠定理论基础,推动 DNA 修饰研究从基础科学领域逐步走向临床应用。
乐备实是国内专注于提供高质量蛋白检测以及组学分析服务的实验服务专家,自2018年成立以来,乐备实不断寻求突破,公司的服务技术平台已扩展到单细胞测序、空间多组学、流式检测、超敏电化学发光、Luminex多因子检测、抗体芯片、PCR Array、ELISA、Elispot、PLA蛋白互作、多色免疫组化、DSP空间多组学等30多个,建立起了一套涵盖基因、蛋白、细胞以及组织水平实验的完整检测体系。
我们可提供从样本运输、储存管理、样本制备、样本检测到检测数据分析的全流程服务。凭借严格的实验室管理流程、标准化实验室操作、原始数据储存体系以及实验项目管理系统,已经为超过3000家客户单位提供服务,年检测样本超过100万,受到了广大客户的信任与支持。








 沪公网安备31011502400759号
沪公网安备31011502400759号
 营业执照(三证合一)
营业执照(三证合一)




