







|
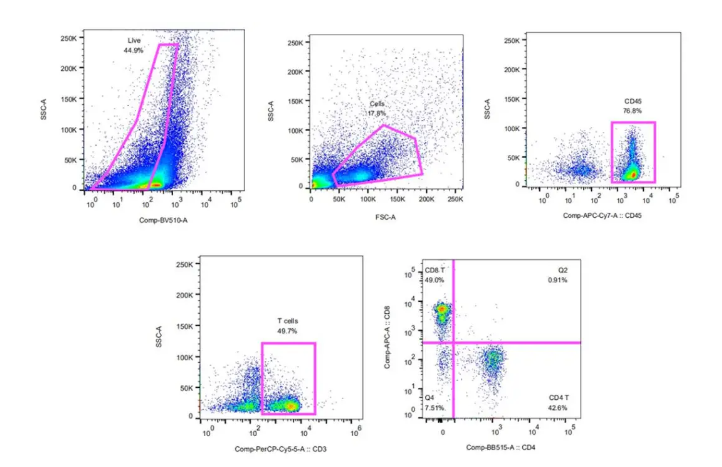
上一期软文中我们对“Immune suppressive landscape in the human esophageal squamous cell carcinoma microenvironment”一文中的图1-4进行了分析,发现在癌症细胞中CD4+、CD8+T细胞,NK细胞与临近组织相比有较明显的变化,且在肿瘤环境中存在免疫抑制。接下来我们进一步针对图5-7进行分析,主要针对巨噬和DC进行了研究。
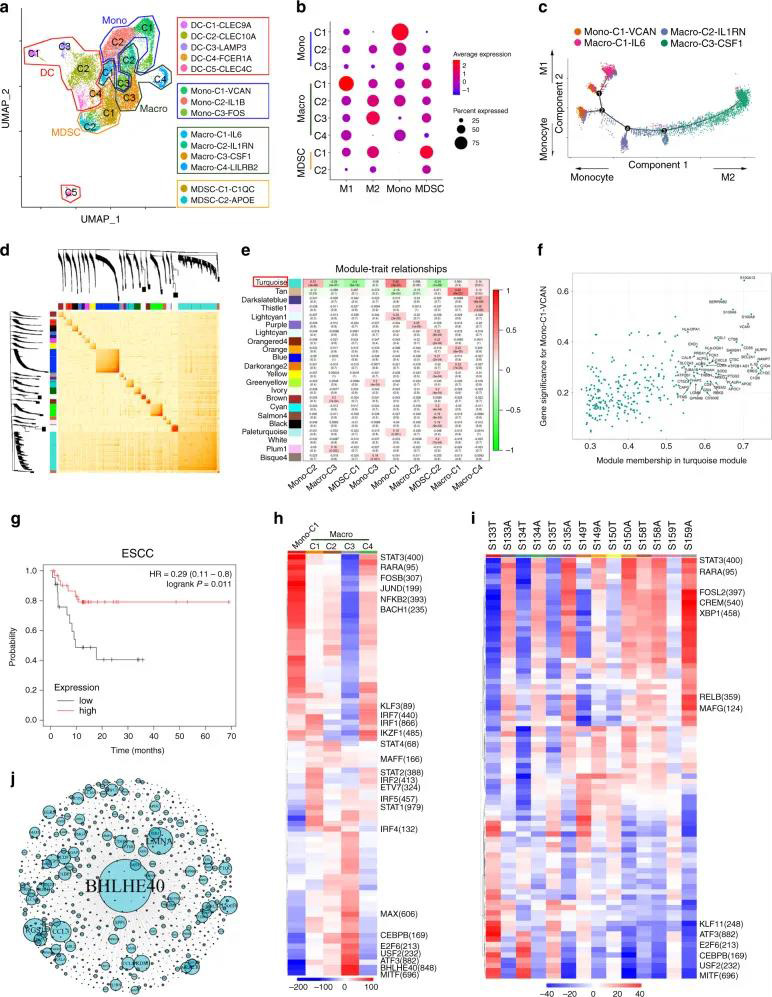
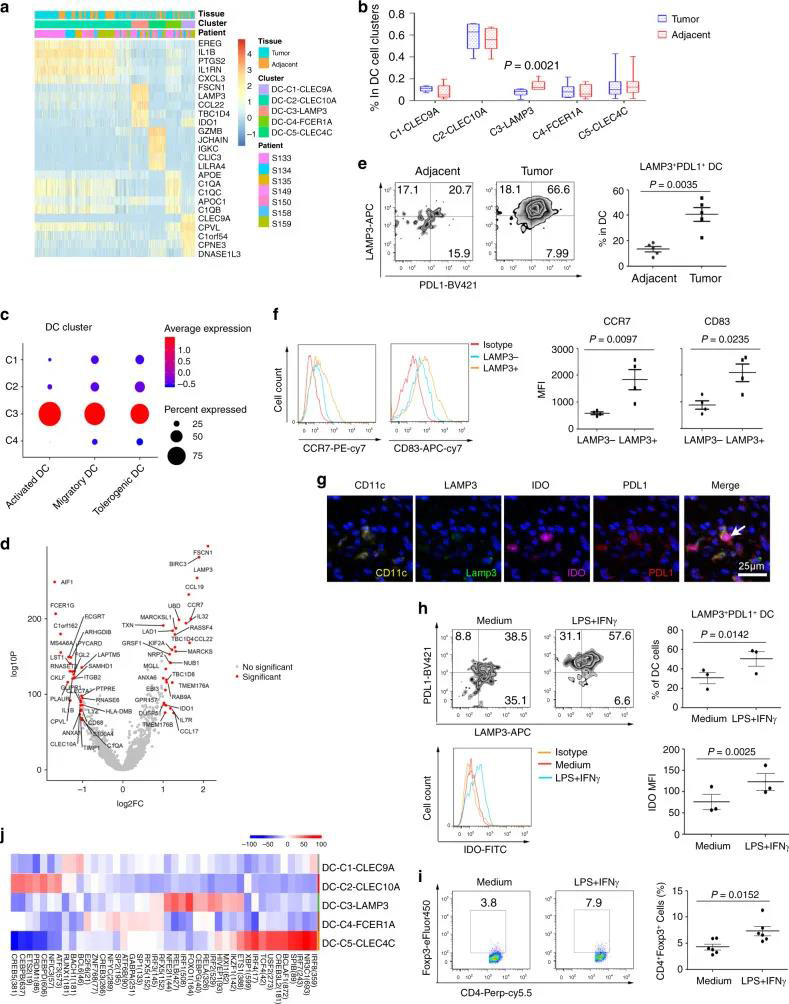
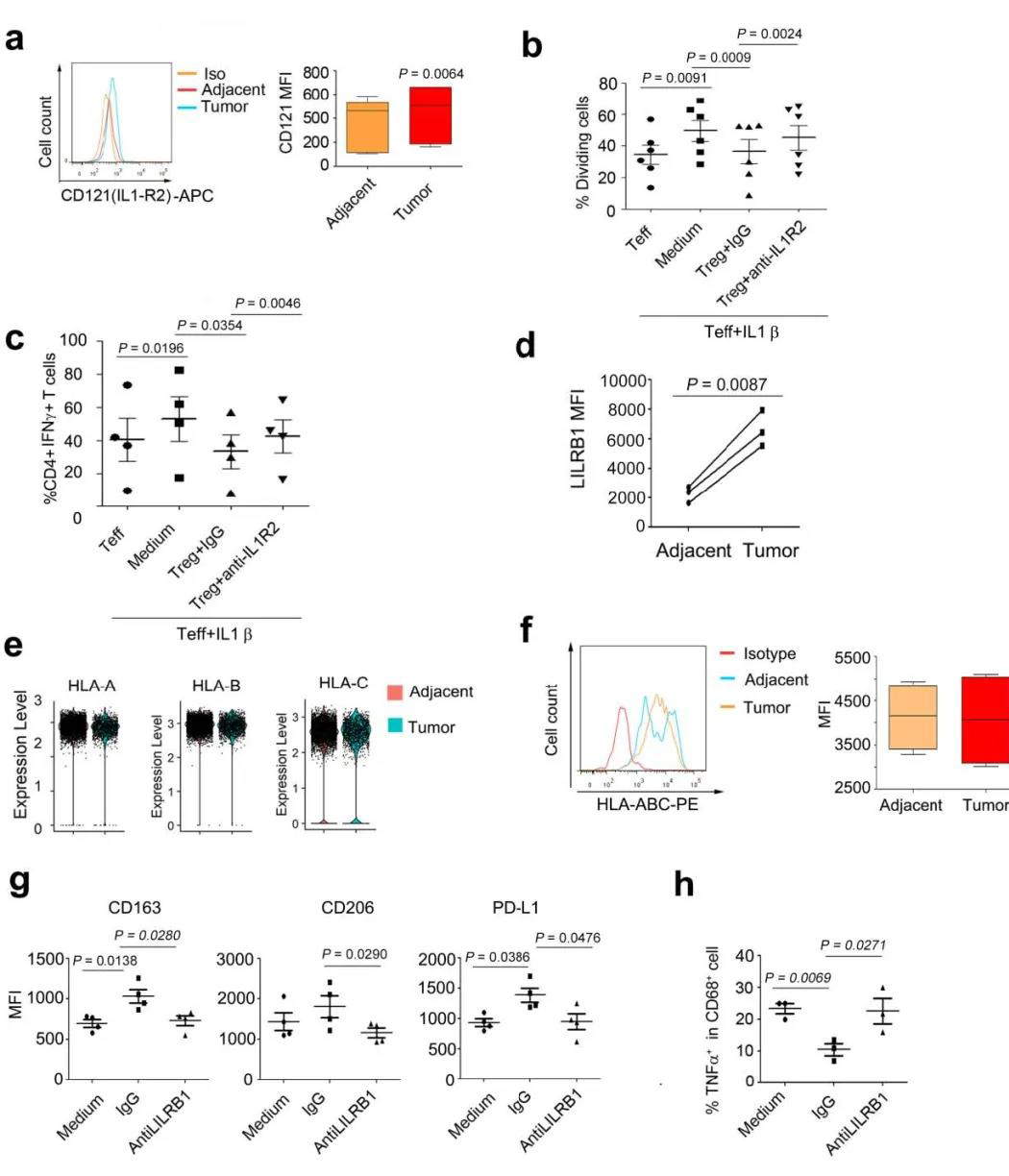
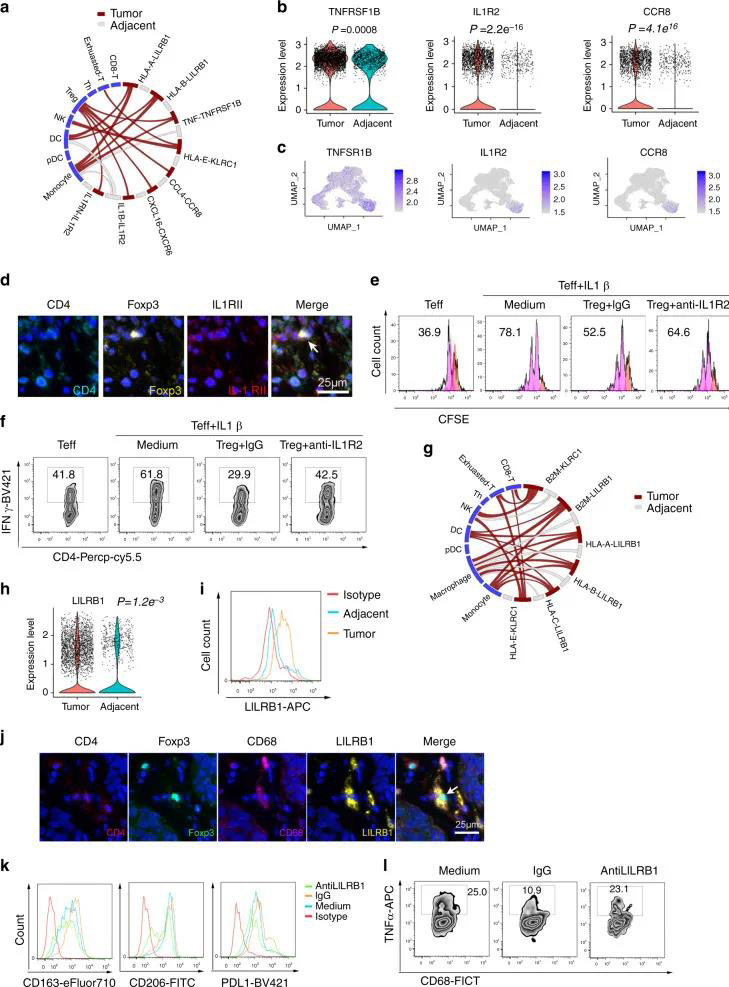
五个DC簇具有高表达水平的CLEC9A,CD1C,FCER1A,LAMP3和CLEC4C,包括常规cDC1(DC-C1-CLEC9A),cDC2(DC-C2-CLEC10A),单核细胞衍生DC(DC-C4-FCER1A) ),LAMP3 + DC(DC-C3-LAMP3)和pDC(DC-C5-CLEC4C)(图6A)。与邻近组织相比,DC-C3-LAMP3富含肿瘤(图6B)。比较了DC的激活和迁移分数,发现LAMP3 + DC与其他DC子集相比具有最高的活性和迁移能力,还发现LAMP3 + DCs丰富了致耐受性的特征(图6C)。LAMP3 + DCs表达了许多调节分子,例如IDO1,EBI3,CD274和IL10(图 6D)。在进行路径富集分析时,发现LAMP3 + DC中上调的基因在细胞因子介导的信号转导,DC细胞分化,白细胞激活,膜运输,抗原加工和呈递的路径中富集(补充图10A),该DC子集支持多功能。 02 、ESCC中免疫细胞间的相互作用
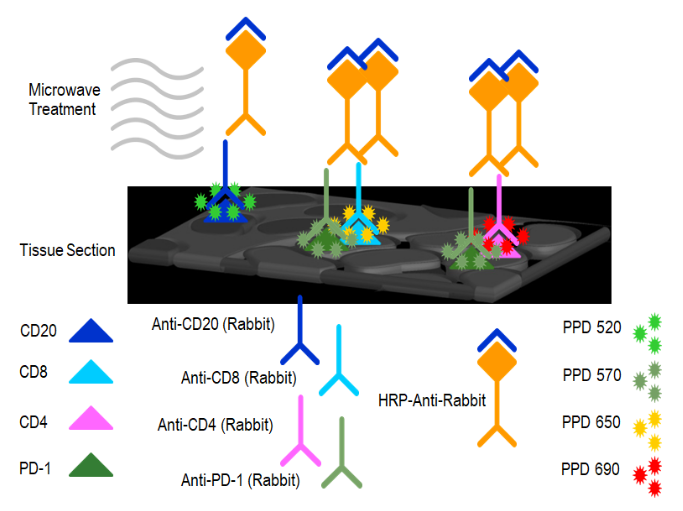
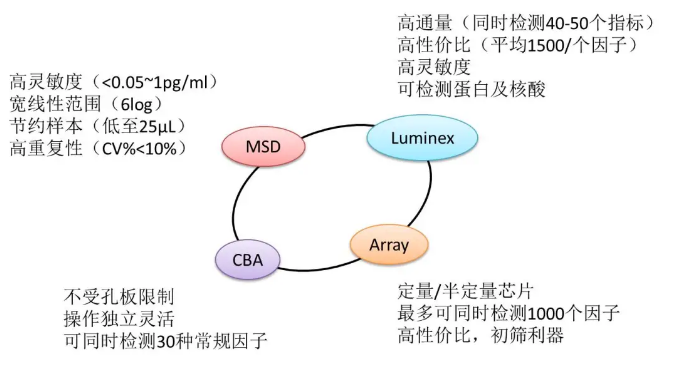
本文涉及技术: 多重免疫组化 本文涉及技术: 单/多因子检测 |









 沪公网安备31011502400759号
沪公网安备31011502400759号
 营业执照(三证合一)
营业执照(三证合一)




