







一、趋化因子CCL11通过调控干性记忆T细胞分化增强核酸疫苗抗肿瘤免疫效应的机制及应用前景
本研究通过大规模体内筛选鉴定出可显著增强核酸疫苗诱导的细胞免疫应答的多种趋化因子。其中,趋化因子CCL19、CCL21及CCL11被证实具有突出的免疫增强作用,尤其以CCL11效果最为显著。进一步机制研究表明,CCL11能够促进抗原特异性干性记忆T细胞的分化,从而显著提升核酸疫苗在肿瘤治疗模型中的免疫保护效果与长期免疫记忆。
本文系统阐释了趋化因子靶向抗原呈递细胞(APC)的疫苗增强策略,该策略不仅可有效提高核酸疫苗激活抗原特异性T细胞的能力,还能显著改善肿瘤微环境中的免疫细胞浸润与功能。值得关注的是,基于该策略优化的mRNA疫苗在多种高负荷肿瘤模型中展现出突出的肿瘤清除效果,为实现实体瘤的免疫治疗提供了新的干预思路。
这一发现为核酸疫苗的免疫增强策略提供了重要的理论依据与实验支持,标志着mRNA肿瘤疫苗在设计与应用方面迈入新的发展阶段,具有重要的转化潜力与研究价值。
二、趋化因子筛选体系的构建与CCL11的功能鉴定
研究团队首先基于成熟的DNA疫苗开发与评价平台,系统构建了包含全部47个人类趋化因子的融合表达文库。每个趋化因子均与模型抗原基因构建为融合表达质粒,并分别免疫小鼠。通过检测抗原特异性T细胞免疫应答水平,初步筛选出6个能显著增强细胞免疫反应的趋化因子候选分子。
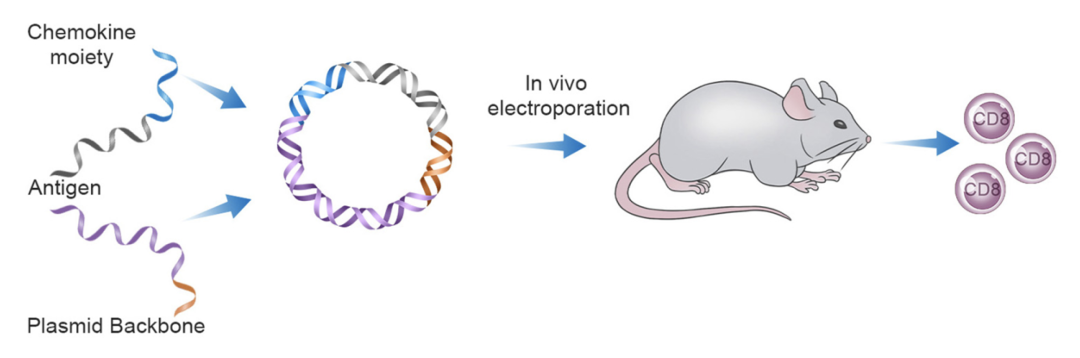 趋化因子筛选模式图
趋化因子筛选模式图
为验证其治疗潜力,进一步在荷瘤小鼠模型中评估了上述6个趋化因子融合疫苗的抗肿瘤效果。实验结果表明,与抗原融合表达的CCL11疫苗展现出最为显著的抗肿瘤活性:在该治疗组中(n=8),所有小鼠的肿瘤均被完全清除,达到100%的肿瘤消退率。同时,生存分析结果显示,CCL11融合疫苗治疗组小鼠的存活率亦为100%。综合上述数据,CCL11被确定为该筛选体系中功能最强的免疫增强趋化因子。
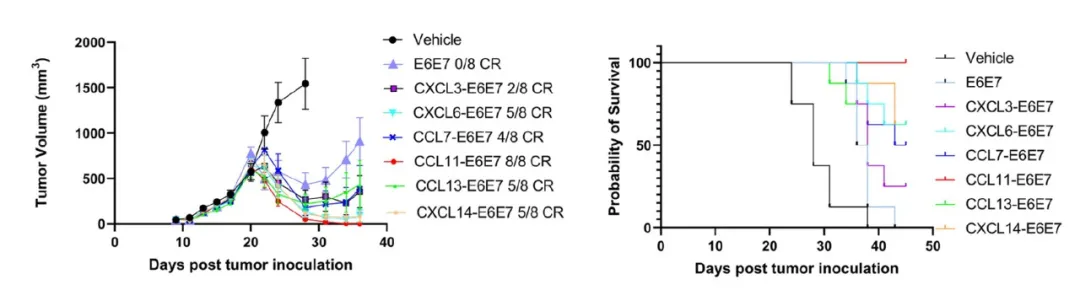 CCL11制备的抗肿瘤疫苗治疗组所有小鼠肿瘤完全清除
CCL11制备的抗肿瘤疫苗治疗组所有小鼠肿瘤完全清除
三、基于趋化因子的mRNA疫苗开发及其免疫机制研究
为深入验证趋化因子-抗原融合策略在不同核酸疫苗平台中的普适性,研究团队将其应用于mRNA疫苗体系。结果显示,该策略能够成功复现DNA疫苗平台中观察到的免疫增强效应,在显著提升抗原特异性T细胞免疫应答的同时,亦有效增强了疫苗的抗肿瘤治疗效果,从而证实其在mRNA疫苗体系中的可行性与有效性。
在此基础上,研究进一步探索了CCL11-抗原融合型mRNA疫苗的潜力。如图所示,相较于溶媒对照组及非融合抗原疫苗组,CCL11-E6E7融合mRNA疫苗在小鼠模型中表现出更为显著的肿瘤生长抑制效果与生存优势,突显了CCL11在增强mRNA疫苗疗效中的关键作用。
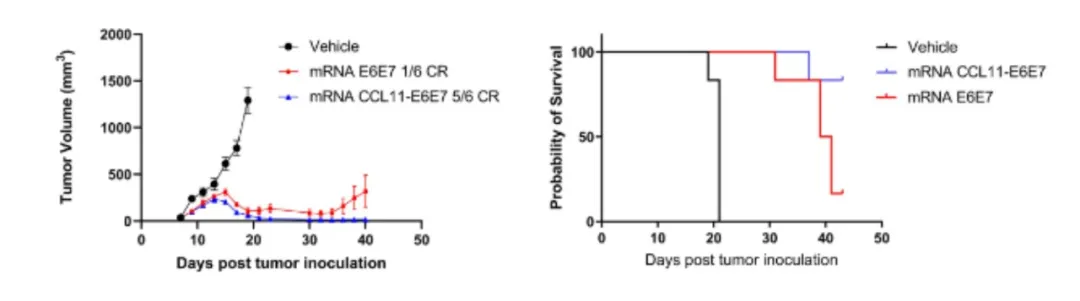 CCL11-E6E7融合型mRNA的抗肿瘤效果优于对照组
CCL11-E6E7融合型mRNA的抗肿瘤效果优于对照组
为进一步提升免疫原性与治疗效率,研究团队基于自复制mRNA(saRNA)技术平台构建了CCL11-E6E7融合疫苗。结果显示,仅需2 μg低剂量免疫即可在已建立约100 mm³肿瘤的小鼠中实现肿瘤的完全清除,不仅体现了saRNA平台的高效抗原表达能力,也进一步证实了趋化因子融合策略在降低有效剂量、提升治疗窗口方面的应用潜力。
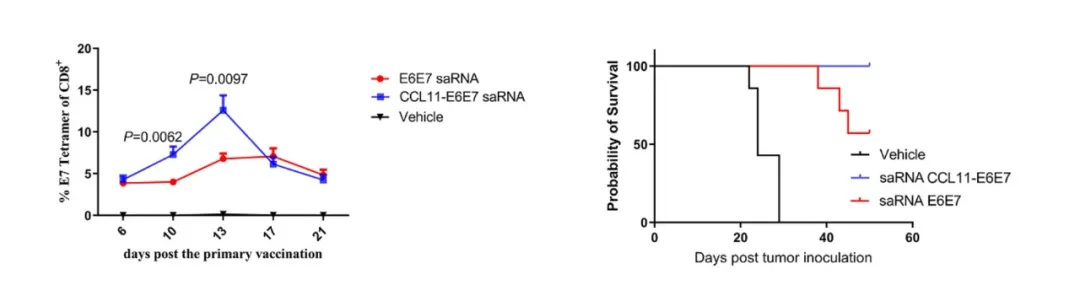 CCL11-E6E7融合型saRNA实现大肿瘤100%清除
CCL11-E6E7融合型saRNA实现大肿瘤100%清除
为深入解析CCL11增强疫苗疗效的免疫学机制,研究团队对治疗后肿瘤微环境进行了分析。结果显示,CCL11疫苗治疗组肿瘤内抗原特异性CD8⁺ T细胞浸润显著增加。RNA-Seq分析进一步表明,该治疗能促进多种免疫细胞向肿瘤部位的聚集,并推动肿瘤微环境向抗肿瘤炎症状态转变(图5),从系统层面揭示了CCL11增强疫苗作用的免疫基础。
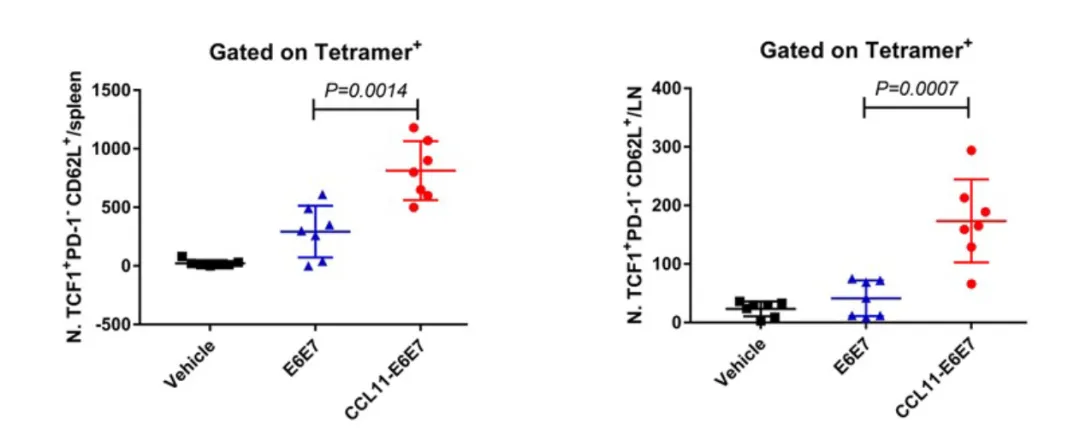
此外,该研究关注了CCL11对记忆T细胞亚群的影响。已有研究提示,淋巴结中的干细胞样记忆T细胞(Tscm)在抗肿瘤免疫中具有重要作用。实验结果表明,相较于对照组,CCL11疫苗免疫后小鼠体内Tscm数量显著增加,说明CCL11能够促进具有长期记忆潜力和强效扩增能力的T细胞分化,这为该策略诱导持久免疫记忆提供了理论依据,也为未来肿瘤免疫治疗方案的优化提供了新的研究方向。
 CCL11制备的疫苗诱导更高质量的特异性T细胞
CCL11制备的疫苗诱导更高质量的特异性T细胞
四、CCL11趋化因子检测服务哪里有?
LabEx为您提供专业、精准的CCL11趋化因子检测服务。作为重要的嗜酸性粒细胞趋化因子,CCL11在过敏反应、寄生虫感染、神经炎症及组织纤维化等多种生理和病理过程中扮演关键角色。
| 货号 | Panel名称 | 种属 | 检测指标 |
| LXLTM36-1 | 小鼠细胞因子/趋化因子-36因子Panel | Mouse | GM-CSF,IFN gamma,IL-1 beta,IL-2,IL-4,IL-5,IL-6,IL-12p70,IL-13,IL-18,TNF alpha,IL-9,IL-10,IL-17A (CTLA-8),IL-22,IL-23,IL-27,G-CSF (CSF-3),IFN alpha,IL-3,IL-15/IL-15R,IL-28,IL-31,IL-1 alpha,LIF,ENA-78 (CXCL5),M-CSF,Eotaxin (CCL11),GRO alpha (CXCL1),IP-10 (CXCL10),MCP-1 (CCL2),MCP-3 (CCL7),MIP-1 alpha (CCL3),MIP-1 beta (CCL4),MIP-2,RANTES (CCL5) |
| LXLTH34-1 | 人细胞因子/趋化因子-34因子Panel | Human | GM-CSF,IFN gamma,IL-1 beta,IL-2,IL-4,IL-5,IL-6,IL-8,IL-12p70,IL-13,IL-18,TNF alpha,IL-9,IL-10,IL-17A (CTLA-8),IL-21,IL-22,IL-23,IL-27,IFN alpha,IL-1 alpha,IL-1RA,IL-7,IL-15,IL-31,TNF beta,Eotaxin (CCL11),GRO alpha (CXCL1),IP-10 (CXCL10),MCP-1 (CCL2),MIP-1 alpha (CCL3),MIP-1 beta (CCL4),RANTES (CCL5),SDF-1 alpha |
| LXLBH27-1 | 人细胞因子-27因子Panel | Human | G-CSF,GM-CSF,IFN-γ,IL-10,IL-12(p70),IL-13,IL-17A,IL-1β,IL-2,IL-4,IL-5,IL-6,IL-7,IL-8/CXCL8,MCP-1/CCL2,MIP-1β,TNF-α,IL-1Rα,IL-9,IL-15,FGF-basic,Eotaxin/CCL11,IP-10/CXCL10,MIP-1α/CCL3,PDGF-BB,RANTES,VEGF-A |
| LXLBH40-1 | 人趋化因子/细胞因子-40因子Panel | Human | 6Ckine/CCL21,BCA-1/CXCL13,CTACK/CCL27,ENA-78/CXCL5,Eotaxin/CCL11,Eotaxin-2/CCL24,Eotaxin-3/CCL26,Fractalkine/CX3CL1,GCP-2/CXCL6,GM-CSF,GRO-α (Gro-a/KC/CXCL1),Gro-β/CXCL2,I-309/CCL1,IFN-γ,IL-1β,IL-2,IL-4,IL-6,IL-8/CXCL8,IL-10,IL-16,IP-10/CXCL10,I-TAC/CXCL11,MCP-1/CCL2,MCP-2/CCL8,MCP-3/CCL7,MCP-4/CCL13,MDC/CCL22,MIF,MIG/CXCL9,MIP-1α/CCL3,MIP-1δ/CCL15,MIP-3α/CCL20,MIP-3β/CCL19,MPIF-1/CCL23,SCYB16/CXCL16,SDF-1α+β/CXCL12,TARC/CCL17,TECK/CCL25,TNF-α |








 沪公网安备31011502400759号
沪公网安备31011502400759号
 营业执照(三证合一)
营业执照(三证合一)




